|
《免疫学和免疫学检验》 > 第三章 补体系统
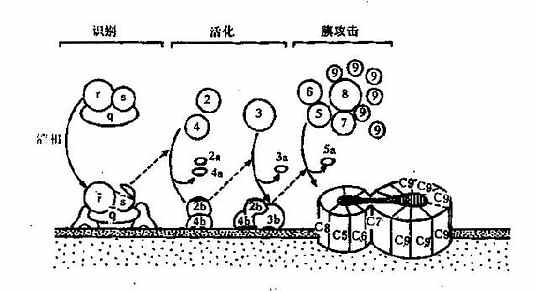
第二节 补体系统的活化与调控补体系统的各组分在体液中通常以非活性状态、类似酶原的形式存在,当受到一定因素激活,才表现出生物活性。补体的激活途径主要有两种,即经典途径(claasicalpathway)和替代途径(alternativepathway)。 (一)经典途径 经典途径是以结合抗原后的IgG或IgM类抗体为主要激活剂,补体C1~C9共11种成分全部参与的激活途径。现发现除抗原抗体复合物外,还有许多因子可激活此途径,如非特异性凝集的Ig、细菌脂多糖、一些RNA肿瘤病毒、双链DNA、胰蛋白酶、纤溶酶、尿酸盐结晶、C-反应蛋白等。经典活化途径可人为地分成识别、活化和膜攻击3个阶段。 1.识别阶段在抗体结合抗原形成复合物后,与C1q结合。IgG1、IgG2、IgG3的补体结合位点在CH2区内,而IgM补体结合位点在CH3区内,IgG4、IgA、IgD和IgE不能结合补体。电镜下观察发现,C1q的球形结构与抗体结合后,进一步激活C1r和C1s,C1s具有酯酶活性,继之进入下一步的连续反应(图3-3)。研究还发现激活C1q的球形分子必须具有2个以上紧密相邻的IgG分子,IgM只需1分子即可,故单分子IgM比IgG激活补体的能力大得多,在补体介导的抗体溶细胞反应中,同量的IgM比IgG更有效。 2.活化阶段此阶段主要形成2种重要的转化酶:C3转化酶C4b3b和C5转化酶C4b2b3b。C4和C2均为C1酯酶的天然底物,C1使C4裂解成C4b和游离的C4a两个片段。C4bα链断端上暴露的硫酯键高度不稳定,可与细胞表面的蛋白质或糖形成共价酰胺键或酯键,在Mg2+存在时C1和C4b一起将C2裂解成大片段C2b和游离的小片段C2a。C2b和C4b结合可形成C4b2b(C3转化酶),将C3裂解成大片段C3b和游离的小片段C3a。继而C3b结合至C4b2b附着的邻近细胞膜上,形成C4b2b3b三分子复合物,即C5转化酶。 3.膜攻击阶段此期形成膜攻击复合物(membraneattackcomplex,MAC)使靶细胞溶解。C5转化酶将C5裂解为C5b和游离的小分子C5a,C5b与细胞膜结合,继而结合C6和C7形成C5b67三分子复合物,C5b67吸附C8,C8是C9的吸附部位,可以与1~18个C9分子结合,并催化C9,使之聚合成内壁亲水的管状跨膜通道,使胞内物质释放出来,水进入细胞,细胞破裂。补体经典途径激活过程见图3-3。
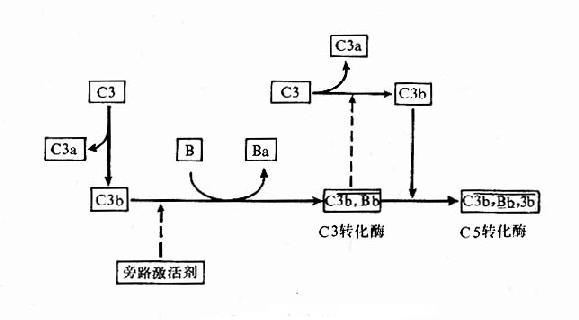
图3-3补体经典激活途径 (二)替代途径 替代途径或称旁路途径,与经典途径的不同之处主要是越过C1、C4和C2,直接激活补体C3,然后完成C5~C9的激活过程;参与此途径的血清成分尚有B、D、P、H、I等因子。替代途径的激活物主要是细胞壁成分,如脂多糖、肽糖苷及酵母多糖等。 1.旁路C3转化酶的形成在生理条件下,血中的C3可受蛋白酶的作用水解少量的C3b,C3b可与邻近的细胞膜结合。如结合的物质是细胞壁上的脂多糖,则C3b的半衰期延长,足以使其与B因子结合形成C3bB复合物。B因子为C3激活剂前体(C3proactivator,C3PA),与结合在膜上的C3b构成C3PA复合物后,使其对D因子的作用更为敏感。D因子为C3PA转化酶原,炎症时增多,在Mg2+存在时转化为活性形式,能使C3bB中的B因子裂解出无活性的小碎片Ba,剩余的C3bBb即旁路C3转化酶。C3bBb与正常血清中活化的P因子(properdin,P)结合成C3bBbP,而使其趋于稳定,减慢衰变。生理条件下C3bBb和C3bBbP使补体系统处于准激活状态,对补体的全面激活具有重要意义。 2.C5激活替代途径的激活物如细菌脂多糖或酵母脂多糖出现时,为C3b和C3bBb提供了可结合的表面,并保护它们不受I因子和H因子的迅速灭活,这时C3激活即由准备状态进入激活状态。C3bBb裂解C3产生C3a和C3b,C3b可与上述的C3bBb,C3bBbP形成多分子的复合物,C3bnBb或C3bnBbP,此即C5转化酶,其作用类似经典途径中的C4b2b3b,可使C5裂解为C5a和C5b,至此以后的补体激活过程与经典途径相同(图3-4)。 3.C3正反馈循环补体活化过程中形成的C3转化酶不断使C3裂解,生成大量的C3b;新产生的C3b又可与B因子结合,扩大进一步的活化,构成了一个正反馈的循环圈,放大了补体的激活作用。不论是经典途径,还是替代途径,只要有C3活化,就可以进入C3正反馈循环,产生放大效应。
图3-4旁路激活途径 (三)补体活化的调控 补体系统被激活后,进行系统有序的级联反应,从而发挥广泛的生物学效应,参与机体的防御功能。但如果补体系统活化失控,可形成过多的膜攻击复合物面产生自身损伤,或过多的炎症介质也会造成病理效应。正常机体的补体活化处于严密的调控之下,从而维持机体的自身稳定。 1.补体的自身调控补体激活过程中生成的某些中间产生非常不稳定,成为补体级联反应的重要自限因素。如C3转化酶C4b2b和C3bBb均易衰变,从而限制了C3的裂解及其后的酶促反应,与细胞膜结合的C4b、C3b及C5b也易衰变,可阻断级联反应。此外,只有细胞表面形成的抗原抗体复合物才能触发经典途径,而旁路途径的C3转化酶则仅在特定的物质表面才具有稳定性,故正常机体内一般不会发生过强的自发性补体激活反应。 2.调节因子的作用体内的存在多种可溶性膜结合的补体调节因子,它们以特定方式与不同的补体成分相互作用,使补体的激活与抑制处于精细的平衡状态,调节蛋白的缺失有时是造成某些疾病发生的原因。目前发现的补体调节蛋白有十余种,按其作用特点可分为三类:①防止或限制补体在液相中自发激活的抑制剂;②抑制或增强补体对底物正常作用的调节剂;③保护机体组织、细胞免遭补体破坏作用的抑制剂。主要的补体调节因子及其功能见表3-3。 表3-3主要补体调节蛋白及其活性
…… 第三节 免疫球蛋白的生物学活性 第四节 各类免疫球蛋白的特点 第五节 免疫球蛋白的基因及抗体形成 第三章 补体系统 第一节 补体系统的组成和性质 第二节 补体系统的活化与调控(当前页) 第三节 补体系统的生物活性 第四节 补体的合成及代谢 第四章 免疫系统 第一节 免疫器官 …… |