|
《人体寄生虫学》 > 第十四章 吸虫
第一节 概论吸虫(trematode)属于扁形动物门的吸虫纲(Class Trematoda)。寄生人体的吸虫属于复殖目(Order Digenea),称为复殖吸虫(digenetic trematode)。复殖吸虫虽然种类繁多,形态各异,生活史复杂,但基本的结构(图14-1)和发育过程略同。
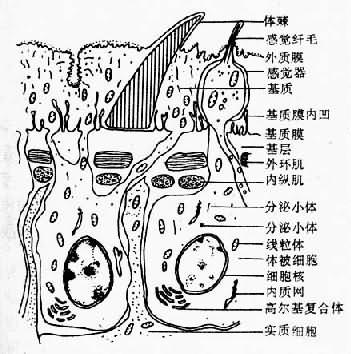
图14-1 复殖吸虫成虫形态构造示意图 形态与功能 大多数的复殖吸虫的成虫外观呈叶状或长舌状,两侧对称,背腹扁平,通常具口吸盘(oral sucker)与腹吸盘(acetabulum),内部结构如下: 体壁组织吸虫成虫体表有凹窝、凸起、皱褶、体棘、感觉乳突等,其形态、数量、分布等随虫种与虫体部位而异。体壁由体被(tegument)与肌肉层组成。中间为实质组织(parenchymal tissue)和埋在实质组织中的消化、生殖、排泄、神经系统等,缺体腔。体被为具有代谢活力的合胞体(syncytium),从外到内由外质膜(external plasma membrane)、基质(matrix)与基质膜(basal plasmamembrane)组成。基质膜之下为基层(basement layer)。肌肉层由外环肌(circular muscle)与内纵肌(longitudinal muscle)组成。体棘位于基质膜之上。感觉器位于基质中,有纤毛伸出体表,另一端有神经突(nerve process)与神经系统相通。此外还有内质网(endoplasmic reticulum)、核糖体(ribosome)、吞噬体(phagosome)、线粒体(mitochondrion)、高尔基复合体(golgi complex)和各种分泌小体(secretory body)等(图14-2)。体被具有保护虫体、吸收营养物质、感觉等生理功能。在结构上,体壁可随虫种、环境条件、发育阶段的不同而有差别,也可出现更新。结构的差别与更新通常也有生理功能上的意义。
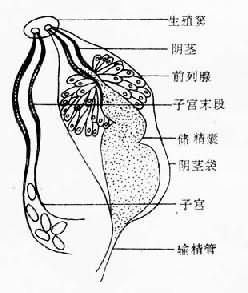
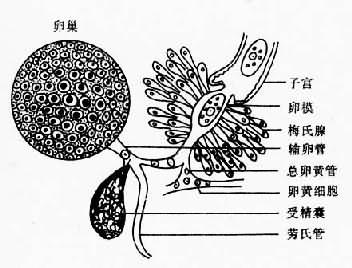
图14-2 复殖吸虫成虫体壁结构示意图 消化系统消化系统由口、前咽(prepharynx)、咽(pharynx)、食管(esophagus)和肠管(intestine)组成。肠管通常分为左右两个肠支(cecum)。肠壁为单细胞层,其外周为肌细胞层所包围,肠细胞含有丰富的粗面内质网与线粒体,还常有高尔基复合体与各种包涵物,其胞质伸出具浆膜的长绒毛样褶,游离于肠腔,以扩大吸收面积。吸虫的消化系统具有消化和吸收功能。消化一般开始于肠管前部而在肠支中完成,主要是细胞外酶的作用。 生殖系统人体吸虫除血吸虫外都具有雌雄两性的生殖器官,称为雌雄同体(hermaphrodite)。雄性生殖系统由睾丸(testis)、输出管(vas efferens)、输精管(vas deferens)、储精囊(seminal vesicle)、前列腺(prostatic gland)、射精管(ejaculatory duct)或阴茎(cirrus)、阴茎袋(cirrus pouch)等组成。雌性生殖由卵巢(ovary)、输卵管(oviduct)、卵模(ootype)、梅氏腺(Mehlis'gland)、受精囊(seminal receptacle)、劳氏管(Laurer's canal)、卵黄腺(vitellaria)、卵黄管(vitelline duct)、总卵黄管(common vitelline duct)、卵黄囊(vitellaria reservior)、子宫(uterus)、子宫末段(metraterm)等组成(图14-3、4)。雌、雄生殖系统的远端在生殖窦(genital atrium)会合,开口
图14-3 复殖吸虫成虫卵巢-卵巢模区结构
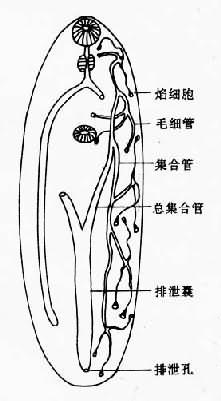
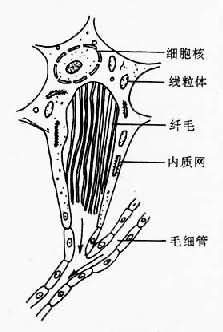
图14-4 复殖吸虫成虫生殖末段结构 于生殖孔(genital pore)。精子产生后从雄性生殖系统转入雌性生殖系统到达受精囊,卵的受精通常在输卵管。受精卵和从卵黄腺来的卵黄细胞排出的卵壳前体物质,在卵模内形成卵壳,以后进入子宫,经生殖孔排出。在复殖吸虫成虫结构系统中,生殖系统最发达,代谢最旺盛。由于成虫每日产卵数量很大,合成代谢与能量代谢都必须满足需要;因此,从外界摄取的营养物质的大部分最终都在本系统消耗。当然,进入虫体的有害物质,包括杀虫药,如无特殊的组织选择性,也同样会大部分积聚在本系统,常造成其正常生理功能首先被损害,表现为产卵数量的下降,甚至完全丧失生殖能力,最后虫体死亡。 排泄系统 吸虫的排泄系统由焰细胞(flame cell)、毛细管(capillary tubule)、集合管(collecting tubule)与排泄囊(excretory bladder)组成,经排泄孔(excretory pore)通体外(图14-5)。焰细胞与毛细管构成原肾(protonephron)单位。焰细胞的数目与排列各种吸虫可有不同,通常用焰细胞式(flame cell pattern)表示。焰细胞(图14-6)有细胞核、线粒体、内质网等;胞浆内有一束纤毛,每一纤毛有两根中央纤丝(fibril)与9根外周纤丝组成。纤毛颤动时很像火焰跳动,因而得名。纤毛的有节律的摆动可带动和保持排泄系统内部液体的流动,并可能形成相当高的过滤压,促使含有氨、尿素、尿酸等废物的排泄液排出虫体外。
图14-5 复殖吸虫成虫排泄系统
图14-6 焰细胞结构 神经系统 咽的两侧各有一神经节(ganglion),有背索(esophageal commissure)相联。神经节向前后各发出三条神经干(nerve trunk),分布于虫体的背面、腹面及侧面。向后神经干之间在不同水平上有横索(transverse commissure)。由神经干发出的神经感觉末梢到达口吸盘、咽、腹吸盘等器官,以及体壁外层的许多感觉器。神经系统有乙酰胆碱酯酶与丁酰胆碱酯酶的活动,神经节中有神经分泌细胞的存在,说明神经系统功能很活跃。 生活史 复殖吸虫的生活史都要经历有性世代(sexual generation)与无性世代(asexual generation)的交替。无性世代一般寄生在软体动物(中间宿主),通常是腹足类(gastropod),如螺蛳等。也可是斧足类(pelecypod),如蚌类。有性世代大多寄生在脊椎动物(终宿主)。复殖吸虫的生活史虽复杂,各种吸虫也有差别,但生活史的基本型则包括卵(ovum)、毛蚴(miracidium)、胞蚴(sporocyst)、雷蚴(redia)、尾蚴(cercaria)、囊蚴(encysted metacercaria)、后尾坳(囊内脱去尾部的虫体称后尾蚴metacercaria)与成虫(adult)。毛蚴侵入螺蛳的淋巴系统内发育变成为胞蚴。胞蚴通过体表摄取营养物质,其体内的胚细胞团经过分裂、发育形成多个雷蚴。雷蚴体内的胚细胞团再分化发育为多个子雷蚴或大量的尾蚴。有的吸虫,在寒冷季节,雷蚴不产生尾蚴而只产生雷蚴,可连续数代。此种变换生殖现象,是多胚繁殖(也称蚴体增殖)的吸虫长期适应环境的结果。绝大多数的尾蚴都有体部和尾部,但形态结构各异。尾蚴从螺体逸出后或侵入第二中间宿主体内或在物体的表面形成囊蚴。有些吸虫缺雷蚴期或有两代以上的雷蚴期,也可缺囊蚴期,而尾蚴直接侵入终宿主发育为成虫。侵入终宿主后童虫通常都经过移行,然后到达定居部位。一般认为移行中的虫体可能识别宿主的不断改变的连续的刺激,使绝大多数的虫体能够按一定的移行途径到达定居部位,而不适宜宿主则不能提供必要的生理信号,因而出现异常的个体发生移行(ontogenetic migration),导致发育迟缓、死亡或异常和异位寄生。复殖吸虫童虫移行时经历的和成虫寄生的器官、系统,其生化环境各异,可以设想必须具备有效的机制和生化代谢方式才能维持生命。各种吸虫从不同的寄生环境摄取的营养物质可能不同,但多以糖原而不以脂类作为能量贮存的一种形式,也缺乏脂类氧化所必需的酶。 生理 人体寄生的吸虫属复殖目,其生活史过程较复杂,既未丧失自生生活的某些特性,又能较广泛适应动物机体内的各种理化条件。这种广泛的适应性和迅速应变能力,是吸虫的重要生理特征之一。 吸虫的营养代谢,一般认为是以糖作为一种重要的能源,蛋白质与脂肪酸较不重要。吸虫吸收已糖主要是通过皮层,以被动扩散或以易化扩散方式进行,后者摄入速度比前者快;吸收后多以糖原形式贮存于虫体实质中。实验证明酵解酶类普遍存在于吸虫,加以许多吸虫的寄生期是处在氧压低的环境中,因此认为无氧糖酵解对吸虫尤为重要;但是糖酵解的整个过程有待进一步研究。氨基酸不是成虫能量主要来源,但胞蚴能分解利用较多的氨基酸。不同吸虫的氨基酸库差别较大,转氨酶种类也不相同,能利用的氨基酸亦各异。吸虫缺少脂类代谢。因而脂类积存于组织和排泄系统中。 吸虫寄居部位的含氧量差别较大,各种吸虫利用氧的途径、需氧的程度各异。氧溶解于吸虫皮层或肠道内壁而进入体内,被游离的血红蛋白携带和储存在各种组织中,复殖目吸虫生活史过程经历几个氧压很不相同的生活环境。例如虫卵、毛蚴和逸离螺体的尾蚴处在氧压高的环境;胞蚴、雷蚴和未成熟的尾蚴处在氧压低的环境;在物体表面结囊的后尾蚴和生活在血液中的成虫处在氧压高的环境,而宿主组织内的囊蚴和消化道中的成虫处在氧压低的环境。因而,在氧压变化的不同环境中生存,吸虫呼吸代谢也发生相应的变化。 分类 我国常见寄生人体的吸虫分类见表14-1。 表14-1 我国常见寄生人体吸虫的分类与其主要寄生部位
…… 第四节 卡氏肺孢子虫 第五节 其它孢子虫 第十三章 纤毛虫 医学蠕虫 第十四章 吸虫 第一节 概论(当前页) 第二节 华支睾吸虫 第三节 布氏姜片虫 第四节 肝片形吸虫 第五节 卫氏并殖吸虫 …… |