|
《生理学》 > 第十章 神经系统
第六节 脑的高级功能和脑电图
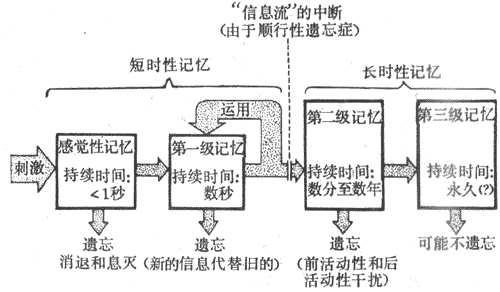
一、学习和记忆学习和记忆是两个相联系的神经过程。学习指人和运动依赖于经验来改变自身行为以适应环境的神经活动过程。记忆则是学习到的信息贮存和“读出”的神经活动过程。 (一)学习的形式1.简单学习简单学习不老大哥在刺激和反应之间形成某种明确的联系,又称为非联合型学习(nonassociative learning)。习惯化和敏感化属于这种类型的学习。习惯化是指当一个不产生伤害性效应的刺激重复作用时,机体对该刺激的反射反应逐渐减弱的过程,例如人们对有规律而重要出现的强噪音逐渐不再对它产生反应。敏感化是指反射反应加强的过程,例如一个弱伤害性刺激本仅引起弱的反应,但在强伤害性刺激作用后弱刺激的反应就明显加强。在这里,强刺激与弱刺激之间并不需要建立什么联系。 2.联合型学习经典条件反射和操作式条件反射均属于联合型学习。 (1)经典条件反射:在动物实验中,给狗吃食物会引起唾液分泌,这是非条件反射。给狗以铃声则不会引起唾液分泌,因为铃声与食物无关,这种情况下铃声称为无关刺激。但是,如果每次给狗吃食物以前先出现一次铃声,然后再给以食物,这样多次结合以后,当铃声一出现,动物就会出现唾液分泌。铃声本来是无关刺激,现在由于多次与食物结合应用,铃声具有了引起唾液分泌的作用,即铃声已成为进食(非条件刺激)的信号。所以这时就把铃声称为信号刺激或条件刺激,这样的反射就称为条件反射。可见,条件反射是在后天生活中形成的。形成条件反射的基本条件就是无关刺激与非条件刺激在时间上的结合,这个过程称为强化。任何无关刺激与非条件刺激结合应用,都可以形成条件反射。 (2)操作式条件反射:操作式条件反射比较复杂,它要求动物完成一定的操作。例如,将大鼠放入实验箱内,当它在走动中偶然踩在杠杆上时,即喂食以强化这一操作;如此重复多次,大鼠即学会了自动踩杠杆而得食。然后,在此基础上进一步训练动物只有当再现某一特定的信号(如灯光)后踩杠杆,才能得到食物的强化。在训练完成后,动物见到特定的信号,就去踩杠杆而得食。这类条件反射的特点是,动物必须通过自己完成某种运动或操作后才能得到强化,所以称为操作式条件反射。 (二)条件反射活动活动的基本规律1.经典条件反射建立需要的基本条件条件反射的建立要求在时间上把某一无关刺激与非条件刺激结合多次,一般条件刺激要先于非条件刺激而出现。条件反射的建立与动物机体的状态有很密切的关系,例如处于饱食状态的运动则很难建立食物性条件反射,动物处于困倦状态也很难建立条件反射。一般来说,任何一个能为机体所感觉的动因均可作为条件刺激,而且在所有的非条件刺激的基础上都可建立条件反射,例如食物性条件反射、防御性条件反射等。 2.经典条件反射的消退条件反射建立之后,如果反复应用条件刺激而不给予非条件刺激强化,条件反射就会逐渐减弱,最后完全不出现。这称为条件反射的消退。例如,铃声与食物多次结合应用,使狗建立了条件反射;然后,反复单独应用铃声而不给予食物(不强化)则铃声引起的唾液分泌量会逐渐减少,最后完全不能引起分泌。巴甫洛夫认为,条件反射的消退是由于在不强化的条件下,原来引起唾液分泌的条件刺激,转化成为引起中枢发生抑制的刺激。从这一观点出发,条件反射的消退并不是条件反射的丧失,而是人原先引起兴奋(有唾液分泌)的条件反射转化为引起抑制(无唾液分泌)的条件反射;前者称为阳性条件反射,后者称为阴性条件反射。 3.人类的条件反射研究动物条件反射的方法,原则上也可用于研究人的条件反射活动;例如,将无关刺激与食物性唾液分泌非条件反射相结合(可用于儿童),或将无关刺激与防御性运动非条件反射相结合等等。此外,人类还可应用词语强化的运动条件反射研究法;例如,当红光在受度儿童面前出现时,实验者说:“按”,受试儿音即用手按压橡皮球。在这一实验中,红光是条件刺激,“按”是词语强化。用词语强化与红光结合2-3次后,如果受试者见到红光信号出现后,立即自动按球,这就形成了对红光的条件反射。 4.两种信号系统学说在人同样可以用光、声、嗅、味、触等感觉刺激作为信号来形成条件反射;这种信号直接作用于眼、耳、鼻、舌、身等感受装置,都是现实具体的信号。此外,抽象的语词也可以代替具体的信号而引起条件反射反应。例如,受试者对每分钟摆动120次的快节拍器声音形成了用温热刺激强化的手臂血管舒张反射,而对每分钟摆动60次的慢节拍器声音形成了用冷刺激强化的血管收缩反射;当这些条件反射被巩固后,实验者对受试者说“快节拍器音”或“慢节拍器音”,这些语词也分别能引起相应有血管舒张或血管收缩反应。如果说具体的信号是第一信号,则相应的语词是第一信号的信号,即第二信号。因此,在人类有两种性质完全不同的信号,第一信号是具体的信号,第二信号(语词)是抽象的信号。巴甫洛夫提出人脑有两个信号系统。第一信号系统是对第一信号发生反应的大脑皮层功能系统,第二信号系统是对第二信号发生反应的大脑皮层功能系统。动物只有一个信号系统,相当于人的第一信号系统;而人类才具有两个信号系统,这是人类区别于动物的主要特征。第二信号系统的发生与发展是人类社会的产物,人类由于社会性劳动与交往产生了语言,语词是现实有概括和抽象化;人类借助于语词来表达其思维,并进行抽象的思维。 (三)记忆的过程外界通过感觉器官进入大脑的信息量是很大的,但估计仅有1%的信息能被较长期地贮存记忆,而大部分却被遗忘。能被长期贮存的信息都是对个体具有重要意义的,而且是反复作用的信息。因此,在信息贮存过程中必然包含着对信息的选择和遗忘两个因素。信息的贮存要经过多个步骤,但简略地可把记忆划分为两个阶段,即短时性记忆和长时性记忆。在短时性记忆中,信息的贮存是不牢固的,例如,对于一个电话号码,当人们刚刚看过但没有通过反复运用而转入长时性记忆的话,很快便会遗忘。但如果通过较长时间的反复运动,则所形成的痕迹将随每一次的使用而加强起来;最后可形成一种非常牢固的记忆,这种记忆不易受干扰而发生障碍。 人类的记忆过程可以细分成四个阶段(图10-43),即感觉性记忆、第一级记忆、第二级记忆和第三级记忆;前二个阶段相当于上述的短时性记忆,后二个阶段相当于长时性记忆。感觉性记忆是指通过感觉系统获得信息后,首先在脑的感觉区内贮存的阶段;这阶段贮存的时间很短,一般不超过1分钟,如果没有经过注意和处理就会很快消失。如果住处在这阶段经过加工处理,把那引起不持续的、先后进来的信息整合成新的连续的印象,就可以从短暂的感觉性记忆转入第一级记忆。这种转移一般可通过两种途径来实现,一种是通过把感觉性高蛋白的资料变成口头表达性的符号(如语言符号)而转移到第一级记忆,这是最常见的;另一种非口头表达性的途径,这在目前还了解得不多,但它必然是幼儿学习所必须采取的途径。但是,信息在第一级记忆中停留的的时间仍然很短暂,平均约几秒钟;通过反复运用学习,信息便在第一级记忆中循环,从布延长了信息在第一级记忆中停留的时间,这样就使信息容易转入第二级记忆之中。第二级记忆是一个大而持久的贮存系统。发生在第二级记忆内的遗忘,似乎是由于先前的或后来的信息的干扰所造成的;这种干扰分别称为前活动干扰和后活动性干扰。有些记忆的痕迹,如自己的名字和每天都在进行操作的手艺等,通过长年累月的运动,是不易遗忘的,这一类记忆是贮存在第三级记忆中的。
图10-43从感觉性记忆至第三级记忆的信息流图解 (四)记忆障碍临床上把记忆障碍分为两类,即顺行性遗忘症(anterogradeamnesia)和逆行性遗忘症(retrograde amnesia)。凡不能保留新近获得的信息的称为顺行性遗忘症。患者对于一个新的感觉性信息虽能作出合适的反应,但只限于该刺激出现时,一旦该刺激物消失,患者在数秒钟就失去作出正确反应的能力。所以患者易忘近事,而远的记忆仍存在。本症多见于慢性酒精中毒者。发生本症的机制,可能是由于信息不能从第一级记忆转入第二级记忆;一般认为,这种障碍与海马的功能损坏有关。前文已述及,海马及其环路的功能遭受破坏,会发生近期记忆障碍。凡正常脑功能发生障碍之前的一段时间内的记忆均已丧失的,称为逆行性遗忘症;患者不能回忆起紧接着本症发生前一段时间的经历。一些非特异性脑疾患(脑震荡、电击等)和麻醉均可引起本症。例如,车祸造成脑震荡的患者在恢复后,不能记起发生车祸前一段时期内的事情,但自己的名字等仍能记得。所以,发生本症的机制可能是第二级记忆发生了紊乱,而第三级记忆却不受影响。 (五)学习和记忆的机制1.从神经生理角度看学习和记忆的机制从神经生理的角度来看,感觉性记忆和第一级记忆主要是神经元生理活动的菜单现。神经元活动具有一定的后作用,在刺激作用过去以后,活动仍存留一定时间,这是记忆的最简单的形式,感觉性记忆的机制可能属于这一类,在神经系统中,神经元之间形成许多环路联系,环路的连续活动也是记忆的一种形式,第一级记忆的机制可能属于这一类。例如,海马环路的活动就与第一级记忆的保持以及第一级记忆转入第二级记忆有关。 近年来对突触传递过程的变化与学习记忆的关系进行了许多研究。在海兔(一种海洋软体动物)的缩鳃反射的研究中观察到,习惯化的发生是由于突触传递出现了改变,突触前末梢的递质释放量减少导致突触后电位减少,从而使反射反应逐渐减弱;敏感化的机制是突触传递效能的增强,突触前末梢的递质释放量增加。在高等动物中也观察到突触传递具有可塑性。有人在麻醉兔中,记录海马齿状回颗粒细胞的电活动观察到,如先以一串电脉冲刺激海马的传入纤维(前穿质纤维),再用单个电刺激来测试颗粒细胞电活动改变,则兴奋性突触后电位和锋电位波幅增大,锋电位的潜伏期缩短。这种易化现象持续时间可长达10小时以上,并被称为长时程增强(long-term potentiation)。不少人把长时程增强与学习记忆联系起来,认为它可能是学习记忆的神经基础。在训练大鼠进行旋转平台的空间分辨学习过程中,记忆能力强的大鼠海马长时程增强反应大,而记忆能力差的大鼠长时程增强反应小。 2.从神经生化角度看学习和记忆的机制从神经生化的角度来看,较长时性的记忆必然与脑内的物质代谢有关,尤其是与脑内蛋白质的合成有关。在金鱼建立条件反射的过程中,如用嘌呤霉素(puromycin)注入动物脑内以抑制脑内蛋白质的合成,则运动不能完成条件反射的建立,学习记忆能力发生明显障碍。人类的第二级记忆可能与这一类机制关系较大。在逆行性遗忘症中,可能就是由于脑内蛋白质合成代谢受到了破坏,以致使前一段时间的记忆丧失。 中枢递质与学习记忆活动也有关。运动学习训练后注射拟胆碱药毒扁豆碱可加强记忆活动,而注射抗胆碱药东莨菪硷可使学习记忆减退。用利血平使脑内儿茶酚胺耗竭,则破坏学习记忆过程。动物在训练后,在脑室内注入γ-氨基丁酸可加速学习。动物训练后将加压素注入海马齿状回可增强记忆,而注入催产素则使记忆减退。一定量的脑啡可使动物学习过程遭受破坏,而纳洛酮可增强记忆。临床研究发现,老年人血液中垂体后叶激素含量减少,用加压素喷鼻可使记忆效率提高;用加压素治疗遗忘症亦收到满意效果。 3.从神经解剖角度看学习和记忆的机制从神经解剖的角度来看,持久性记忆可能与新的突触联系的建立有关。动物实验中观察到,生活在复杂环境中的大鼠,其大脑皮层的厚度大,而生活在简单环境中的大鼠,其大脑皮层的厚度小;说明学习记忆活动多的大鼠,其大脑皮层发达,突触的联系多。人类的第三级记忆的机制可能属于这一类。
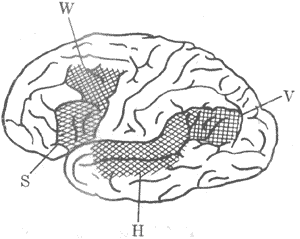
二、大脑皮层的语言中枢和一侧优势(一)两侧大脑皮层功能的相关两侧大脑皮层之间有许多连合纤维,在哺乳类动物中最大的连合纤维结构是胼胝体;进化愈高等则胼胝体愈发达,人类的胼胝体估计含有100万根纤维。有人观察到,当在犬的身体一侧皮肤上给予刺激,并与食物或酸防御性唾液分泌反射相结合形成条件反射后,则另一侧皮肤相应部位的机械刺激也自然具有阳性的条件反射效应。如果事先将该动物的胼胝体切断,则这种现象就不能出现。还有人事先切断猫视交叉的交叉纤维,使一侧眼睛的视网膜传入冲动仅向同侧皮层投射,然后将动物一眼蒙蔽,用另正好学习对图案的鉴别能力;待其学会后将该眼蒙蔽,测定先前被蒙蔽眼的图案鉴别能力,见到先前被蒙蔽的眼也能具有这种鉴别能力。如果事先切断这个动物的胼胝体,则这种现象就不能出现。由此看来,两侧大脑皮层的感觉分析功能是相关的,胼胝体连合纤维能将一侧皮层的活动向另一侧转送。电生理研究指出,刺激对侧皮层对应点可以加强这一侧皮层的感觉传入冲动的诱发电们,起着易化作用。这一易化作用是通过胼胝体连合纤维完成的,因为这类纤维主要联系两侧皮层相对应的部位。在人类,两侧大脑皮层的功能也是相关的,两关球之间的连合纤维对完成双侧的运动、一般感觉和视觉的功调功能有重要作用。右手学会了一种技巧运动,左手虽然没有经过训练,但在一定程度上也会完成这种技巧运动,说明一侧皮层的学习活动可以通过连合纤维向另一侧转送。 (二)大脑皮层的语言中枢人类大脑皮层一定区域的损伤,可以引致特有的各种语言活动功能障碍(图10-44)。临床发现,损伤布洛卡(Broca)三角区(44区,在中央前回底部之前,图中S区),会引致运动失语症(motor aphasia)。病人可以看懂文字与听懂别人谈话,但自己却不会讲话,不能用语词来口头表达;然而,其与发音有关的肌肉并不麻痹,就是不能用“词”来表达自己的意思。损伤额中回后部接近中央前回手部代表区的部位(图中W区),则病人可以听懂别人的谈话,看懂文字,自己也会讲话,但不会书写;然而,其手部的其他运动并不受影响,这种情况称为失写症(agraphia)。颞上回后部(图中H区)的损伤,会引致感觉失语症(sensory aphasia),病人可以讲话及书写,也能看懂文字,但听不懂别人的谈话;事实上,病人能听到别人的发音,就是不懂其含义,但其视觉却是良好的,其他的语言活动功能仍健全,这种情况称为失读症(alexia),因此,语言活动的完整功能是与广大皮层区域的活动有关的,各区域的功能是密切相关的。严重的失语症可同时出现上述四种语言活动功能的障碍。
图10-44 人大脑皮层语言功能的区域 V区障碍不能认识词义 H区障碍不能听懂话 S区障碍不能讲话 W区障碍不能书写 (三)大脑皮层功能的一侧优势产生上述各种语言活动功能障碍时,在一般运用右手劳动为主的成年人中,其大脑皮层损伤经常发生在左侧。因为绝大多数用右手劳动为主的成年人,右侧大脑皮层的44区的损伤并不发生明显的语言活动障碍;然而其左侧大脑皮层布洛卡的损伤,则可形成严重的运动失语症,这种左侧大脑皮层的语言活动功能上占优势的现象,反映了人类两侧大脑半球功能是不对等的,这种一侧优势的现象仅在人类中具有。 人类左侧大脑皮层在语言活动功能上占优势的现象,虽然与一定的遗传因素有关,但主要是在后天生活实践中逐步形成的,这与人类习惯运用右手进行劳动有密切的关系。小儿在2-3岁之前,如果发生左侧大脑半球损害时,其语言活动功能的紊乱和右侧大脑半球损害时的情况没有明显的差别;说明这时候尚未建立左侧优势,双侧大脑半班干部均与语言活动功能有关。10-12岁时,左侧优势逐步建立;但在左侧大脑半球损害后,尚有可能在右侧大脑半球损害就很难在右侧大脑皮层再建立起语言活动的中枢。在发育成年人后,左侧优势已经形成,如果发生左侧大脑半球损害就很难在右侧大脑皮层再建立起语言活动的中枢。在运用左手劳动为主的人中,则左右双侧的皮层有关区域都可能成为语言活动的中枢。有人进行过统计,在48例运用右手劳动的人中,语言中枢在左侧的为43例,在右侧的为25例,4例左右双侧均有关;在20例左右手混用的人中,语言中枢在左侧的12例,在右侧的为2例,6例左右双侧均有关。 由于左侧大脑半球在语言活动功能上占优势,因此一般称左侧半球为优势半球或主要半球,右侧半球为次要半球。但是研究指出,右侧半球也有其特殊的重要功能。目前知道,右侧大脑皮层在非语词性的认识功能上是占优势的,例如对于空间的辩认、深度知觉、触觉认识、音乐欣赏分辨等等。右侧大脑皮层顶叶损伤的病人,由于非语词性认识能力的障碍,常再现穿衣失用症(apraxia);患者虽然没有肌肉麻痹,但穿衣困难,他会将衬衣前后穿倒或只将一只胳膊伸入袖内。右侧大脑皮层顶叶、枕叶、颞叶结合处损伤的病人,常分不清左右侧,穿衣困难,不以维持绘制图表。右侧大脑半球后部的病变,常发生视觉认识障碍;患者不能辨认别人的面部,甚至不能认识镜子里自己的面部,而且还伴有对颜色、物体、地方的认识障碍。 上述两侧大脑半球对不同认识功能的优势现象,还可通过裂脑(splitbrain)实验研究加以证实。在患有顽固性癫痫发作的病人,为了控制癫痫在两半球之间传布发作,常将患者的连合纤维(胼胝体)切断;手术后患者对出现在左侧视野中的物体(视觉投射到右侧半球)不能用语词说出物体的名称,而对出现在右侧视野中的物体(视觉投射到左侧半球)就以臁出物体的名称,说明语言活动中枢在左侧半球。但是,患者右侧半球的视觉认识功能是良好的。譬如,先给患者的左侧视野看一支香烟,他不能用语词说出这一物体是“香烟”;但是患者认识到这一物体是香烟,因为他可以闭着眼睛藉助于触觉有矩手把许多香烟收集起来以表示他对这一物体的认识。在正常人,虽然语言活动中枢在左侧半球,但能对左侧视野中的物体说出其名称,这是连合纤维的功能,因为连合纤维使左右两侧半球的功能发生了联系。 一侧优势是指人类的脑的高级功能向一侧半球集中的现象;左侧半球在语词活动功能上占优势,右侧半球在非语词性认识功能止占优势。但是,这种优势敢是相对的,而不是绝对的;因为左而半球也有一定的非语词性认识功能,右侧半球也有一定的简单的语词活动功能。
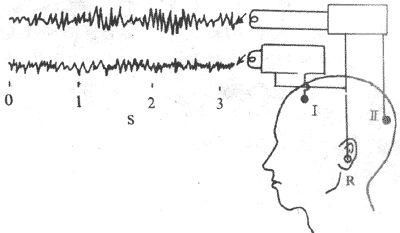
三、脑电图和脑诱发电位大脑皮层的神经元具有生物电活动,因此大脑皮层经常有持续的节律性电位改变,称为自发脑电活动。临床上在头皮用双极或单极记录法来观察皮层的电位变化,记录到的脑电波称为脑电图(图10-45)。在动物中将颅骨打开或以病人进行脑外科手术时,直接在皮层表面引导的电位变化,称为皮层电图。此外,在感觉传入冲动的激发下,脑的某一区域可以产较为局限的电位变化,称为脑诱发电位。
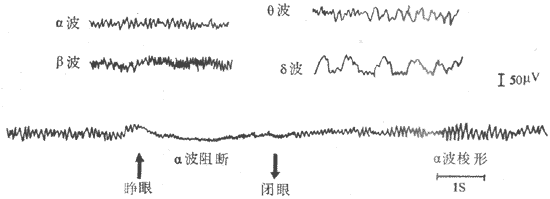
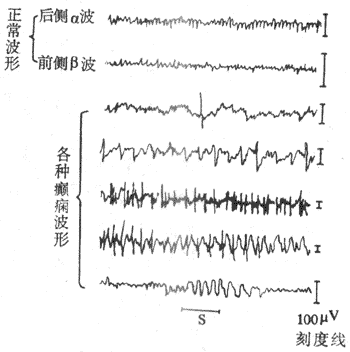
图10-45 脑电图记录示意图 无关电极放置在耳壳(R),由额叶(I)电极导出的脑电波振幅低, 由枕叶(Ⅱ)导出的脑电波振幅高频率较慢 (一)脑电图的波形脑电图的波形分类,主要是依据其频率的不同来人工划分的。在不同条件下,波形频率的快慢可有显着的差别,每秒0.5-3次的波称为δ波,4-7次的波称为θ波,8-13次的波称为α波,14-30次的波称为β波。一般说来,频率慢的波其波幅常比较大,而频率快的波其波幅就比较小。例如,在成年人头上皮上引导时,δ波可有20-200μV,α波有20-100μV,而β波只有5-20μV(图10-46)。
图10-46 正常脑电波各种波形 各种波都可在皮层的不同区域引得,但枕叶区域其α波活动比较显著,而β波在额叶与顶叶比较显著。有时,β波与α波同时在一个部位出现,而β波重合在α波的上面。人类α波在清醒、安静并闭眼时即出现。α波出现时,在枕叶部位最大,并可具有时大时小的变化;即波幅先由小逐渐变大,然后又由右面变小,接着又由小变大,如此反复,形成α波的梭形,每一梭形持续约1-2s。睁开眼睛或接受其他刺激时,α波立即消失而呈现快波,这一现象称为α波阻断,如果被试者又安静闭眼时,则α波又重现。在困倦时,一般可见θ波。成人清醒状态下,几乎是没有θ波的,但在睡眠期间皮层脑电图可出现δ波。如将睡者唤醒,δ波即转成快波。因此,一肌认为快波是新皮层处在紧张活动状态时的主要脑电活动表现,α波是皮层处在安静状态时的主要表现,慢波是睡眠状态下的主要表现。在幼儿时期,脑电波频率比成人慢,一般常见到θ波,到10岁后才出现明确的α波;在婴儿时期,脑电波频率更慢,常见到δ波。此外δ波在成年人极度疲劳时及麻醉状态下也可出现。 癫痫患者,脑电图可出现棘波、尖波、棘慢综合波等,棘波的时程在80ms以下,幅度为50-150μV。尖波的时程为80-200ms,幅度为100-200μV。棘慢综合波指的是棘波后跟随出现一个慢波,慢波时程达200-500ms;一般棘慢综合波出现时,多数为每秒3次左右(图10-47)。在皮层具有占位性病变(肿瘤等)的区域,即使病人外于清醒状态时,亦可引出θ波或δ波。因此,临床上可以借这些脑电波改变的特点,并结合临床资料,来诊断癫痫或探索肿瘤的所在部位。
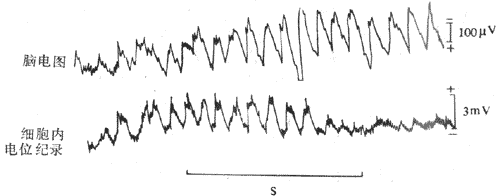
图10-47脑电图正常波形与癫痫波形的对比 (二)脑电波形成的机制脑电波以的波形是一种近似于正弦波的电位变化,而与神经干上见到的动作电位不一样。应用微电极记录皮层神经元细胞内电位变化,见到皮层表面出现类似α波节律的电位变化时,细胞内记录到的突触后电位变化也出现节律相一致的改变(图10-48)。由此认为,,此层表现的电位变化主要是由突触后电位变化形成的,也就是说由细胞体和树突的电位变化形成的。可以设想,单一神经元的突触后电位变化是不足以引起皮层表面的电位改变的;必须有大量的神经组织同时发生突触后电位变化,才能同步起来引起皮层表面再现电位改变。从皮层的神经元组成来看,锥体细胞的分布排列比较整齐,其顶树突互相平行并垂直于皮层表面,因此其电活动在同步时易于总和而形成强大的电场,从而改变皮层表面的电位。
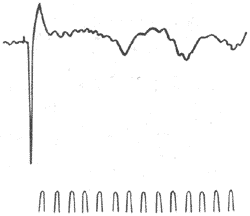
图10-48脑电图与皮层神经元细胞内电位记录的关系 目前知道,大量皮层神经组织的放电活动同步总和必须依赖丘脑的功能。在动物实验中见到,当用中度麻醉时,即使没有其他感觉传入的刺激,皮层会出现每称8-12次的自发脑电活动。这种脑电活动的波幅亦时大时小,并可以皮层广泛的区域内引出,因此这咱脑电活动与人类脑电波中的α节律极相似。如果切断皮层与丘脑间的纤维联系,上述类似α波的电活动就大大减小。如用每秒8-12次节律性电刺激来刺激丘脑非特异投射系统的一此神经核(如髓板内核群),则皮层上会出现每称8-12次的节律性脑电变化。这种变化的波幅亦时大时小,同时在皮层的空间分布也是广泛的,因此,从频率、波幅形状以及空间分布上来看,刺激丘脑非特投射系统所获得的脑电变化,与上述类似α波的自发脑电活动相一致。由此认为,某引起自发脑电形成的同步机制,就是皮层与丘脑非特异投射系统之间的交互作用;一定的同步节律的丘脑非特特异投射系统的活动,促进了电层电活动的同步化。 如果用每秒60次的节律性电刺激来刺激丘脑非特异投射系统,则皮层上类似α波的自发脑电活动立即消失而转成快波。这可理解为高频刺激对同步化活动的扰乱,脑电出现了去同步化现象,快波的出现就是去同步化的结果。刺激脑干网状结构时引起的上行激动作用,一般也认为是其上行冲动扰乱了丘脑非特异投射系统与皮层之间同步化环节,脑电出现了激活状态,呈现了去同步化的快波。在人类脑电记录中所见到的α波阻断现象,事实上也是由同样机制引起的。 电生理研究观察到,当皮层癫痫病灶区出现棘波时,皮层内神经元出现爆发式短串冲动发放,频率可高达每秒200-900次;如将电极插入神经元细胞体内,则观察到当棘波出现时,细胞体出现大幅度去极化电位(可达30mV),去极化电位发展到一定程度后则爆发短串动作电位。由此认为,许多神经元同时出现大幅度的去极化电位,就使皮层表现出现电棘波;而神经元的爆发式短串冲动发放,也是由于大幅度去极化电位造成的,这种大幅度去极化电位,可能是大量同步的兴奋性突触后电位总和起来形成的,这是癫痫病例灶区神经元异常活动的表现。由于皮层肿瘤等占位性病变区本身不产生电活动变化,因此在该区域记录到的θ波或δ波,一般认为来源于其环周异常脑细胞的电活动。 (三)脑诱发电位诱发电位是指感觉传入系统受刺激时,在中枢神经系统内引起的电位变化。受刺激的部位可以是感觉器官、感觉神经或感觉传导途径上的任何一点。但是广义地说,用其他刺激方法引起的中枢神经系统的电位变化,也可称为诱发电位。例如,直接刺激脊髓前根,冲动沿运动神经逆向传至脊髓前多角引起的电位变化,亦可称为诱发电位。 大脑皮层诱发电位一般是指感觉传入系统受刺激时,在皮层上某一局限区域引出的电位变化;由于皮层随时在活动着并产生自发脑电波,因此诱发电位时常出现在自发脑电波的背景之上。在动物皮层相应的感觉区表面引起的诱发电位可分为两部分,一为主反应,另一为后发放(图10-49)。主反应出现的潜伏期是稳定不变的,为先正后负的电位变化。后发放尾随主反应之后,为一系列正相的周期电位变化。皮层诱发电位是用以寻找感觉投射部位的重要方法,在研究皮层功能定位方面起着重要的作用。
图10-49家兔大脑皮层感觉运动区诱发电位 上线:诱发电位记录,向下为正,向上为负 下线:时间,50ms第一个向上小波为刺激桡浅神经记号, 间隔10ms后即出现先正后负的主反应,再 间隔100ms左右后,即相继出现正相波动的后发放 诱发电位也可在人体头颅外头皮上记录到。由于记录电极离中枢较远,颅骨的电阻很大,记录到的电位变化极微弱;而且诱发电位夹杂在自发脑电之间,电位很难分辨。运用电子计算机将电位变化叠加、平均起来,能够使诱发电位显示出来,这种方法记录到的电位称为平均诱发电位(averaged evoked potential)。平均诱发电位目前已成为研究人类的感觉功能、神经系统疾病、行为和心理活动的一种手段。临床常用的有体感诱发电位、听觉诱发电位和视觉诱发电位几种。现简述体感诱发电位的引导方法和波形;刺激电极安放在上肢正中神经经过的皮肤表面(也可放在下肢的某一部位),记录电极放在颅顶靠近中央后回的头皮表面,参考电极置于耳壳;记录到的标准波形如图10-50所示。图中的P9波起源于正中神经的第一级神经元;P11波可能起源于脑干或颈脊髓,因为丘脑以上中枢病变时,P11不受影响,而颈脊髓病变时P11消失;P13和P14波可能由脑干内侧丘系活动所产生;N20波是一个负波,一般认为它来源于丘脑向皮层的投射或皮层感觉区,因为在丘脑病变时可使N20波消失,而N20波以前的电波成分不受影响。因此,通过体感诱发电位的记录和分析,有助于对患者中枢损伤位置的诊断。
四、觉醒和睡眠觉醒和睡眠都是生理活动所必要的过程,只有在觉醒状态下,人体才能进行劳动和其他活动;而通过睡眠,可以使人体的精力和体力得到恢复,于睡眠后保持良好的觉醒状态。成年人一般每天需要7-9小时,儿童需要睡眠的时间比成年人长,而老年需要睡眠的时间就比较短。与觉醒对比,睡眠时许多生理功能发生了变化,一般表现为:①嗅、视、听、触等感觉功能暂时减退;②骨骼肌反射运动和肌紧张减弱;③伴有一系列自主神经功能的改变。例如,血压下降、心率减慢、瞳孔缩小、尿量减少、体温下降、代谢率减低、呼吸变慢、胃液分泌可增多而唾液分泌减少、民汗功能增强等。 (一)觉醒状态的维持动物实验中观察到,单纯在中脑肉状结构的头端加以破坏,而保留各种感觉上传的特异传导途径,动物即进入持久的昏睡状态;各种感觉刺激都不能唤醒动物,脑电波不能由同步化慢波转化成去同步化快波,虽然这时感觉传入冲动完全可以沿特异传导途径抵达大脑皮层。因此认为,觉醒状态的维持是脑干网状结构上行激动系统的作用。目前认为,脑干网状结构上行激动系统可能是乙酰胆碱递质系统,因此静脉注射阿托品能阻断脑干网状结构对脑电的唤醒作用。 进一步的研究观察到,动物在注入阿托品后,脑电呈现同步化慢波而不再出现快波,但动物在行为上并不表现睡眠。看来,觉醒状态的纤维比较复杂,脑电觉醒状态(呈现快波)与行为觉醒状态的维持有不同的机制。动物实验观察到,单纯破坏中脑黑质多巴胺递质系统后,则动物在行为上不能表现觉醒,对新异的刺激不能表现控究行为,但脑电仍可有快波出现。因此,行为觉醒的维持可能是黑质多巴胺递质系统的功能。动物实验还见到,破坏蓝斑上部(去甲肾上腺素递质系统)后,则动物电波快波明显减少;但如有感觉刺激传入冲动时,则动物仍能唤醒脑电呈现快波,不过和种唤醒作用很短暂,感觉传入刺激一停止,唤醒作用即终止。所以,蓝斑上部去甲肾上腺素递质系统与脑电觉醒的维持也有关系。其作用是持续的紧张性作用;而上行激动系统(乙酰胆碱递质系统)的作用是时相性作用,它调制去甲肾上腺素递质系统的脑电觉醒作用。 (二)睡眠的时相通过对整个睡眠过程的仔细观察,发现睡眠具有两种不同的时相状态。其一是脑电波呈现同步化慢波的时相,其二是脑电波呈现去同步化的时相。前者是一般熟知的状态,其表现已在前文述及,常称为慢波睡眠(slow wave sleep,SWW)。后者的表现与慢波睡眠不同,称为异相睡眠(paradoxicalsleep,PS)或快波睡眠、快速眼球运动 (rapid eyemovements,REM)睡眠。异相睡眠期间,各种感觉功能进一步减退,以致锅醒阈提高;骨骼肌反射运动和肌紧张进一步减弱,肌肉几乎完全松驰;脑电波呈现去同步纶快波。这些表现是异相睡眠期间的基本表现。此外,在异相睡眠期间还会有间断性的阵发性表现,例如眼球出现快速运动、部分躯体抽动,在人类还观察到血压升高和心率加快,呼吸加书本而不规则。 慢波睡眠与异相睡眠是两个相互转化的时相。成年人睡眠一开始首先进入慢波睡眠,慢波睡眠持续约80-120分钟左右后,转入异相睡眠;异相睡眠持续约20-30分钟左右后,又转入慢波睡眠;以后又转入异相睡眠。整个睡眠期间,这种反复转化约4-5次,越接近睡眠后期,异相睡眠持续时间逐步延长。在成年人,慢波睡眠和异相睡眠均可直接转为觉醒状态;但觉醒状态只能进入慢波睡眠。而不能直接进入异相睡眠。在异相睡眠期间,如将其唤醒,被试者往往会报告他正在做梦。据统计,在191例被试者异相睡眠期间唤醒后,报告正在做梦的有152例,占80%左右;在160例被试者慢波睡眠期间唤醒后,报告正在做梦的只有11例,占7%左右。因此一般认为,做梦是异相睡眠的特征之一。 在人体中还观察到,垂体前叶生长激素的分泌与睡眠的不同时相有关。在觉醒状态下,生长激素分泌较小;进入慢波睡眠后,生长激素分泌明显升高;转入异相睡眠后,生长激素分泌又减少。看来,慢波睡眠对促进生长、促进体力恢复是有利的。 异相睡眠是睡眠过程中再现的生理现象,具有一定的生理意义。曾观察到,如几天内被试者在睡眠过程中一出现异相睡眠就将其唤醒,使异相睡眠及时阻断,则被试者会出现易激动等心理活动的扰乱。然后,又让被试者能自然睡眠而不予唤醒,开始几天异相睡眠增加,以补偿前阶段异相睡眠的不足;在这种情况下异相睡眠可直接出现在觉醒之后,而不需经过慢波睡眠阶段。由此认为异相睡眠是政党衙所必需的生理活动过程。动物脑灌流实验观察到,异相睡眠期间脑内蛋白质合成加快。因此认为,异相睡眠对于幼儿神经系统的成熟有密切关系;并认为异想睡眠期间有昨于建立新的突触联系而促进学习记忆活动。看来,异相睡眠对促进精力的恢复是有利的。但是,异相睡眠会出现间断性的阵发性表现,这可能与某些疾病在夜间发作有关,例如心绞痛、哮喘、阻塞性肺气肿缺氧发作等。有人报导,病人在夜间心绞痛发作前常先做梦,梦中情绪激动,伴有呼吸加快、血压升高、心率加快,以致心绞痛发作而觉醒。 (三)睡眠发生的机制睡眠是由中枢内发生了一个主动过程而造成的,中枢内存在着产生睡眠的中枢,有人认为,在脑干尾端存在能引起睡眠和脑电波同步化的中枢。这一中枢向上传导可作用于大脑皮层(有人称之为上行抑制系统),并与上行激动系统的作用相对抗,从而调节着睡眠与觉醒的相互转化。 由于中枢神经递质研究的进展,已把睡眠的发生机制与不同的中枢递质系统功能联系了起来。慢波睡眠可能与脑干内5-羟色递质系统有关,异相睡眠可能与脑干内5-羟色胺和去甲肾上腺素递质系统有关。 (张镜如) 参考资料 1.徐丰彦,张镜如主编.人体生理学.北京:人民卫生出版社,1989 2.韩济生主编.神经科学纲要.北京:北京医科大学、中国协和医科大学联合出版社1993 3.潘家右,伍嘉宁. NO:中枢神经中一个新的重要信使物质.生理科学进展1993;24:293-297 4.包雅琳,郭 恒怡.M受体亚型研究的新进展。生理科学进展 1992;23:18-22 5.钱家庆.a –肾上腺素受体,生理科学进展1988;19:57-60 6.王崇铨,突触前受体,生理科学进展1989;20:144-147 7.宋亮年,中枢神经系统中受体亚型的多样性及其生物学意义。生理科学进展1992;23:13;17 8.柳川,神经生长因子的研究,生理科学进展1990;21:54-57 9.邱一华,王健鹤,神经递质和神经肽对免疫功能的影响,生理科学进展1992;22:21-24 10.苏正昌,等,免疫反应产物对神经及内分泌系统功能的影响,生理科学进展1992;23:327-331 11.金观源,突触的可塑性与学习、记忆机制,生理科学进展1989;20:148-153 12.BaileyCH,Kandel ER.Structural changes accompanying memory storage.Ann ReyPhysiol1993;55:397-426 13.Bredt DS.Snyder SH.Nitricoxide,a noval neuronal messenger,Neuron 1992;8:3-11 14.Goodman LS,Gilman A.ThePharmacological Basis of Therapeutics 8thed,Pergamon Press,NewYork,1990 15.Hellige JB.Hemisphericasymmetry.Ann Rev Psychol 1990;41:55-80 16.McNaughton BL.The mechanism ofexpression of long-term enhancement of hippocampal synpses.Ann Rev Physiol1993;55:375-369 17.Ochs S. Systems of materialtransport in nerve fibres (axoplasmic transport )related to nerve function andtrophic control.Ann NY Acad Sci 1974;228:202-223 18.Patton HD,Fuchs AF,HilleB,Scher AM,Steiner R.Textbook of Physiology 21st ed,WB SaunderCO,Philadelphia,1989 19.Turek FW.Circadian neuralrhythms in mammals.Ann Rev Physiol 1985;47:49-64 20. Walsh K.Neuropsychology,Churchill Livingstone,Edinburgh,1978y
…… 第一节 神经元活动的一般规律 第二节 反射活动的一般规律 第三节 神经系统的感觉分析功能 第四节 神经系统对躯体运动的调节 第五节 神经系统对内脏活动的调节 第六节 脑的高级功能和脑电图(当前页) 第十一章 内分泌 第一节 概述 第二节 下丘脑的内分泌功能 第三节 垂体 …… |