|
《生物化学与分子生物学》 > 第十三章 结缔组织生化(Biochemistryof Connective Tissue)
第二节 胶原蛋白胶原蛋白(collagen)存在所有多细胞动物体内,是体内含量最多的一类蛋白质,存在于几乎所有组织中,是一种细胞外蛋白质,以不溶纤维形式存在,具高度抗张能力,是决定结缔组织韧性的主要因素。
一、胶原蛋白的结构单个的Ⅰ型胶原分子分子量约285kD,宽14A,长约3000A。由三条多肽链组成。哺乳动物个体中有30种不同的多肽链构成16种不同的胶原,其中最常见的列于表13-3中。 表13-3 胶原的主要类型
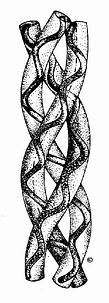
图13-6 胶原的右手超螺旋结构 胶原蛋白的氨基酸组成有如下特征:①甘氨酸几乎占总氨基酸残基的三分之一,即每隔两个其他氨基酸残基(X,Y)即有一个甘氨酸,故其肽链可用(甘-X-Y)n来表示。②含有较多在其他蛋白质中少见的羧脯氨酸和羧赖氨酸残基,也有较多脯氨酸(pro)和赖氨酸。如脯氨酸(Pro)和4—羟脯氨酸(4—hydroxyproline,Hyp)含量高达15~30%。同时还含有少量3—羟脯氨酸(3hydroxyproline)和5—羟赖氨酸(5—hydroxylysine,Hyl)。羟脯氨酸残基可通过形成分子内氢键稳定胶原蛋白分子。例如,正常胶原在39℃变性,而在缺乏脯氨酸羟化酶条件下合成的胶原在24℃变性成为白明胶(gelatin)。而羟赖氨酸上可结合半乳糖-葡萄糖苷,与特定组织功能相关。如在基底膜胶原(Ⅳ型)中含hyl较多,含糖也较多,可能与基底膜的滤过功能有关。③胶原中缺乏色氨酸,所以它在营养上为不完全蛋白质。 在胶原纤维中,胶原蛋白分子单位称为原胶原(tropo collagen)。每个原胶原分子由三条α-肽链组成,α-肽链自身为α螺旋结构,三条α-肽链则以平行、右手螺旋形式缠绕成“草绳状”三股螺旋结构(图13-6)。肽链中每三个氨基酸残基中就有一个要经过此三股螺旋中央区,而此处空间十分狭窄,只有甘氨酸适合于此位置,由此可解释其氨基酸组成中每隔两个氨基酸残基出现一个甘氨酸的特点。而且三条α-肽链是交错排列的,因而使三条α-肽链中的Gly、X、Y残基位于同一水平上,借Gly中的N-H基与相邻链X残基上羟基形成牢固的氢键(图13-7),以稳定其分子结构。 原胶原分子平行排列成束,通过共价交联,可形成稳定的胶原微纤维(microfibvil),进一步行聚集成束,形成胶原纤维。胶原分子通过分子内或分子间的交联成为不溶性的纤维。因胶原分子氨基酸组成中缺乏半胱氨酸,不可能象角蛋白那样以二硫键相联,而是通过组氨酸与赖氨酸间的共价交联,一般发生在胶原分子的C-或N末端之间。
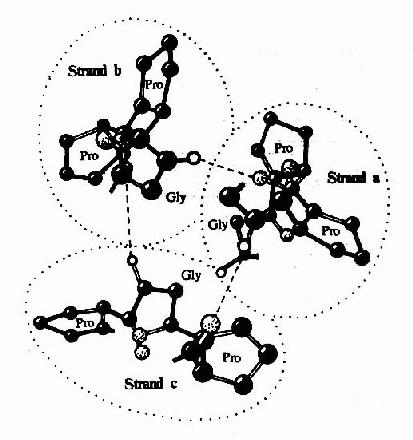
图13-7 胶原样多聚体三螺旋轴的C端投影图 胶原纤维在不同组织中的排列方式与其功能相关。如在肌腱、皮肤及软骨,要分别在一维、二维和三维方向承受张力,因而其胶原纤维排列分别为平行束状,多角的纤维片层及不规则排列等方式。(表13-4) 表13-4 胶原纤维在不同组织中的排列
二、胶原的生物合成结缔组织中的原胶原分子主要由成纤维细胞合成,软骨中胶原由软骨细胞合成,骨胶原来自成骨细胞,基底膜中胶原则由上皮或内皮细胞合成。胶原的生物合成可分为细胞内和细胞外两大阶段。
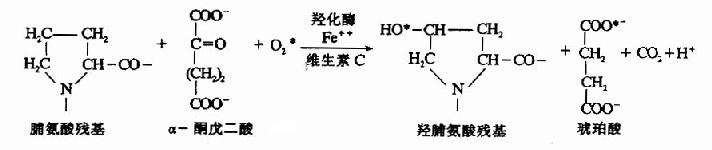
图13-8 胶原结构示意图 1.细胞内合成阶段在结缔组织细胞中,首先是按蛋白质合成的原则先合成一条很长的,约1400个氨基酸残基的肽链,称为溶胶原蛋白,而后转入内质网中进行羟基化和糖基化修饰。 (1)羟基化修饰 胶原分子组成中含有羟脯氨酸和羟赖氨酸,这两种氨基酸并无遗传密码、反密码及tRNA引导入肽链,而是在内质网中,由脯氨酸、赖氨酸残基经羟化生成的。由脯氨酸羟化酶(prolylhydroxylase)和赖氨酸羟化酶(lysyl hydroxylase)催化,此酶为加单氧酶,需Fe++和维生素C为辅因子,α-酮戊二酸作辅底物。如脯氨酸的羟化反应: 此羟化反应中需分子氧,缺氧会妨碍胶原的生成,因而缺氧可使伤口愈合延迟。而维生素C是维持羟化酶活性所必需的,缺乏维生素C,胶原合成不能形成正常的纤维,可出现皮肤结节,血管脆弱及伤口愈合缓慢等症状。羟化作用对三股螺旋的坚固性有重要作用,羟化不足的链在体温下不能形成坚固的三股螺旋,因而不能从细胞内排出。
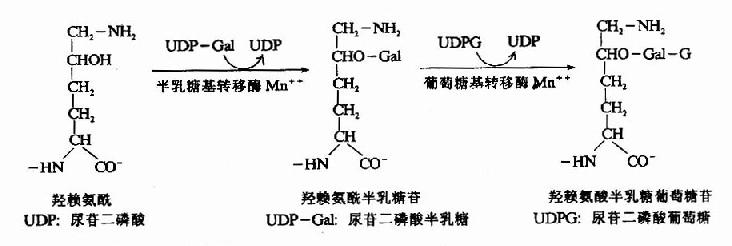
(2)糖基化修饰 胶原分子中含有共价连接的糖基,根据组织不同,糖含量可达0.4-12%。其中糖基主要为葡萄糖、半乳糖及它们的双糖。在内质网中由半乳糖基转移酶及葡萄糖基转移酶催化将糖基联于5-羟赖氨酸残基上。
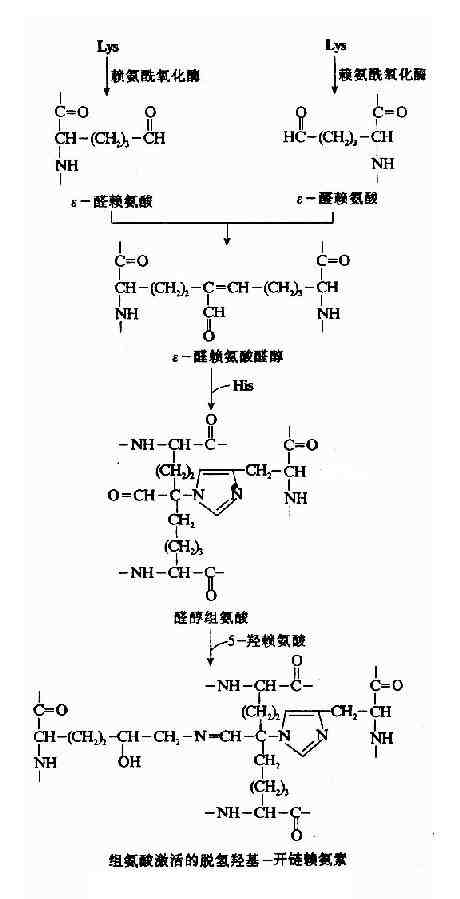
糖基化的作用目前尚未完全阐明,研究发现这些糖基位于胶原纤维中原胶原的接头处。推测糖基化与纤维的定向排列有关。 经羟基化和糖基化修饰的溶胶原蛋白,形成三股螺旋而排出细胞外。 2.细胞外胶原纤维成熟阶段分泌到细胞外的溶胶原由内切酶作用,水解N—末端和C—末端的附加肽链,形成原胶原蛋白,原胶原分子可在中性pH条件下,借分子间各部分不同电荷的相互吸引而自动聚合成胶原纤维,此种聚合不稳定,经共价交联成网使之进一步固定。 胶原纤维的共价交联由赖氨酸氧化酶(lysyloxidase)催化,此酶含Cu++,是参与交联反应过程的唯一酶,能将赖氨酸转变为醛赖氨酸(allysine)ε-醛赖氨酸与另一α-肽链的ε赖氨酸醛醛缩合生成ε-醛赖氨酸醛酸(Allysinealdol),再与组氨酸反应生成醇醛组氨酸(aldol histidine),后者再与5-羟赖氨酸进行醛胺缩合形成希夫碱结构(图13-9)可使4条α-肽链间共价交联。 通过共价交联,胶原微纤维的张力加强,韧性增大,溶解度降低,最终形成不溶性的胶原纤维。
图13-9 胶原纤维的交联过程 胶原蛋白分子共价交联对胶原正常功能有重要意义。例如人或动物进食山藜豆所致的山藜豆中毒(lathyrivsm)表现为严重的骨、关节及大血管壁的异常,其发病即因为山藜豆毒素β—氨基丙晴(β—mino propionitrile)
与赖氨酸氧化酶活性位点共价结合而使之失活,从而导致胶原纤维变脆弱。 胶原蛋白与许多人类疾病相关。如骨形成不全(osteogenesis imperfecta)或称脆骨病(brittle bonediesease),即是由于胶原蛋白生物合成或转录后的修饰作用障碍所致。现已清楚是由于I型胶原突变所致,据突变发生的位置及性质不同,发病的严重性亦不同。此外,有些胶原性疾病涉及多种胶原合成紊乱或合成过程中酶(如赖氨酸羟化酶,赖氨酸氧化酶)活性的异常。如EhlersDanlos综合症即有10种以上不同胶原的缺陷,其症状主要为关节过度伸张,皮肤弹性增高、脆弱,血管脆弱及外伤后皮下粘蛋白或皮下脂肪小结形成等,又被称为“印第安橡皮人”(Indiarubber man)。
三、胶原的分解代谢胶原纤维由于广泛的共价交联,其结构稳定,不易被一般蛋白酶水解。体内有特异作用于胶原的胶原酶(calagenase),对其分解起关键作用。此酶在N椂嗽?/4处切断原胶原,切断后的碎片可自动变性,经细胞外非特异性蛋白酶及肽酶水解或被细胞吞噬后由溶酶体酶进一步分解,形成小分子寡肽或游离氨基酸。 胶原酶在某些修复或再生组织如分娩后子宫、重建的骨组织以及愈合的伤口等含量较高,Ca++为其激活剂。血清中α2巨球蛋白能与胶原酶结合而抑制其活性。胶原酶对温度十分敏感,36℃时酶活性比30℃大10倍。炎症局部温度升高,可能因此加速胶原分解。 胶原分解后产生的寡肽及氨基酸中均含有羟脯氨酸,这在其它蛋白分解产物中少见。因而测定尿中羟脯氨酸或含羟脯氨酸的短肽的含量,可作为判断体内胶原分解的指标。
|