|
《生物化学与分子生物学》 > 第十八章 蛋白质的生物合成(The Biosynthesis of protein)
第三节 蛋白质合成后的分泌及加工修饰不论是原核还是真核生物,在细胞浆内合成的蛋白质需定位于细胞特定的区域,有些蛋白质合成后要分泌到细胞外,这些蛋白质叫做分必蛋白。在细菌细胞内起作用的蛋白质一般靠扩散作用而分布到它们的目的地。如内膜含有参与能量代谢和营养物质转运的蛋白质;外膜含有促进离子和营养物质进入细胞的蛋白质;在内膜与外膜之间的间隙称为周质,其中含有各种水解酶以及营养物质结合蛋白。 真核生物细胞结构更为复杂,而且有多种不同的细胞器,它们又具有各不相同的膜结构,因此合成好的蛋白质还要面临跨越不同的膜而到达细胞器械,有些蛋白质在翻译完成后还要经过多种共价修饰,这个过程叫做翻译后处理。
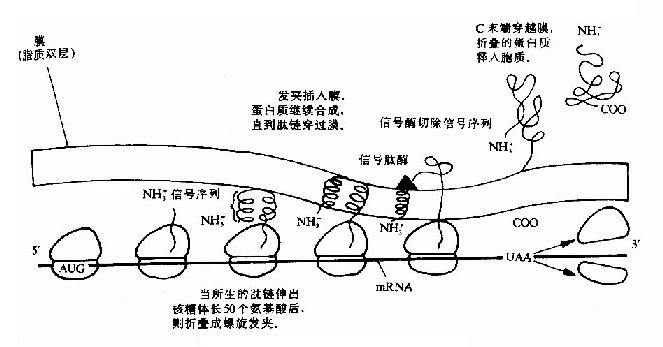
(一)细菌中蛋白质的越膜细胞的内膜蛋白,外膜蛋白和周质蛋白是怎样越过内膜而到其目的地的呢?绝大多数越膜蛋白的N端都具有大约15-30个以疏水氨基酸为主的N端信号序列或称信号肽。信号肽的疏水段能形成一段α螺旋结构。在信号序列之后的一段氨基酸残基也能形成一段α螺旋,两段α螺旋以反平行方式组成一个发夹结构,很容易进入内膜的脂双层结构,一旦分泌蛋白质的N端锚在膜内,后续合成的其它肽段部分将顺利通过膜。疏水性信号肽对于新生肽链跨膜及把它固定的膜上起一个拐掍作用。之后位于内膜外表面的信号肽酶将信号肽序列切除。当蛋白质全部翻译出来后,羧端穿过内膜,在周质中折叠成蛋白质的最终构象(图18-18)。
图18-18 蛋白质合成后的分泌过程
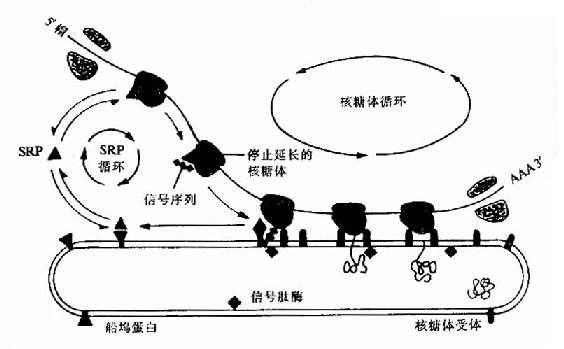
(二)真核生物蛋白质的分泌真核生物不但有细胞核、细胞质和细胞膜,而且还有许多膜性结构的细胞器,在细胞须内合成的蛋白质怎样的到达细胞的不同部位呢?了解比较清楚的是分泌性蛋白质的转运。 像原核细胞一要,真核细胞合成的蛋白质N端也有信号肽也能形成两个α螺旋的发夹结构,这个结构可插入到内质网的膜中,将正在合成中的多肽链带和内质网内腔。80年代中期在胞浆中发现一种由小分子RNA和蛋白质共同组成的复合物,它能特异地与信号肽识别而命名为信号肽识别颗粒。它的作用是识别信号肽与核糖体结合并暂时阻断多肽链的合成。内质网外膜上的SRP受体,当ARP与受体结合后,信号肽就可插入内质网进入内腔,被内质网内膜壁上的信号肽酶水解除去SRP与受体结合后,信号肽就可插入内质网进入内腔,被内质网内腔壁上的信号肽酶水解除去SRP与受体解离并进入新的循环,而信号肽后序肽段也进入内质网内腔,并开始继续合成多肽链(图18-19)。
图18-19 在蛋白质越过内质网的转运过程中,SRP和船坞蛋白(或SRP受体)的作用 SRP对翻译阶段作用的重要生理意义在于:分泌性蛋白及早进入细胞的膜性细胞,能够正确的折叠、进行必要的后期加工与修饰并顺利分泌出细胞。 现以哺乳动物的胰岛素为例说明这种分泌过程。胰岛素由51个氨基酸残基组成,但胰岛素mRNA的翻译产和在兔网织红细胞无细胞翻译体系中为86个氨基酸残基,称为胰岛素原,在麦胚无细胞翻译系统中为110个氨基酸残基组成的前胰岛素原,后来证明,在前胰岛素原的N末端有一段富含疏水氨基酸的肽段做为信号肽,使前胰岛素原能穿越内质网膜进入内质网内腔,在内腔壁上信号肽被水介。所以在哺乳动物细胞内,当多肽链合成完成时,前胰岛素原已成为胰岛素原。然后胰岛素原被运到高尔基复合体,切去C肽成为成熟的胰岛素,最终排出胞外。像真核细胞的前清蛋白,免疫球白轻链,催乳素等都有相似的分必方式。
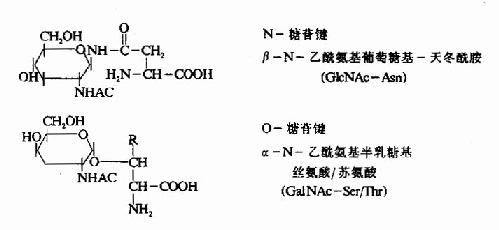
(三)蛋白质翻译后加工修饰从核糖体上释放出来的多肽链,按照一级结构中氨基酸侧链的性质,自竹卷曲,形成一定的空间结构,过去一直认为,蛋白质空间结构的形成靠是其一级结构决定的,不需要另外的信息。近些年来发现许多细胞内蛋白质正确装配都需要一类称做“分了伴娘”的蛋白质帮助才能完成,这一概念的提出并未否定“氨基酸顺序决定蛋白空间结构”这一原则。而是对这一理论的补充,分子伴娘这一类蛋白质能介导其它蛋白质正确装配成有功能活性的空间结构,而它本身并不参与最终装配产物的组成。目前认为“分子伴娘”蛋白有两类,第一类是一些酶,例如蛋白质二硫键异构酶可以识别和水解非正确配对的二硫键,使它们在正确的半胱氨酸残基位置上重新形成二硫键,第二类是一些蛋白质分子,它们可以和部分折叠或没有折叠的蛋白质分子结合,稳定它们的构象,免遭其它酶的水解或都促进蛋白质折叠成正确的空间结构。总之“分子伴娘”蛋白质合成后折叠成正确空间结构中起重要作用,对于大多数蛋白质来说多肽链翻译后还要进行下列不同方式的加工修饰才具有生理功能。 1.氨基端和羧基端的修饰 在原核生物中几乎所有蛋白质都是从N-甲酰蛋氨酸开始,真核生物从蛋氨酸开始。甲酰基经酶水介而除去,蛋氨酸或者氨基端的一些氨基酸残基常由氨肽酶催化而水介除去。包括除去信号肽序列。因此,成熟的蛋白质分子N-端没有甲酰基,或没有蛋氨酸。同时,某些蛋白质分子氨基端要进行乙酰化在羧基端也要进行修饰。 2.共价修饰 许多的蛋白质可以进行不同的类型化学基团的共价修饰,修饰后可以表现为激活状态,也可以表现为失活状态。 (1)磷酸化: 磷酸化多发生在多肽链丝氨酸,苏氨酸的羟基上,偶尔也发生在酪氨酸残基上,这种磷酸化的过程受细胞内一种蛋白激酶催化,磷酸化后的蛋白质可以增加或降低它们的活性,例如:促进糖原分解的磷酸化酶,无活性的磷酸化酶b经磷酸化以后,变居有活性的磷酸化酶a。而有活性的糖原合成酶I经磷酸化以后变成无活性的糖原合成酶D,共同调节糖元的合成与分介。 (2)糖基化: 质膜蛋白质和许多分泌性蛋白质都具有糖链,这些寡糖链结合在丝氨酸或苏氨酸的羟基上,例如红细胞膜上的ABO血型决定簇。也可以与天门冬酰胺连接。这些寡糖链是在内质网或高尔基氏体中加入的(图18-20)。
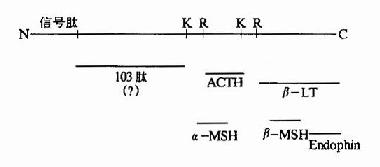
图18-20 糖蛋白中常见的糖一肽连接键 (3)羟基化: 胶原蛋白前α链上的脯氨酸和赖氨酸残基在内质网中受羟化酶、分子氧和维生素C作用产生羟脯氨酸和羟赖氨酸,如果此过程受障碍胶原纤维不能进行交联,极大地降低了它的张力强度。 (4)二硫键的形成: mRNA上没有胱氨酸的密码子,多肽链中的二硫键,是在肽链合成后,通过二个半胱氨酸的疏基氧化而形成的,二硫键的形成对于许多酶和蛋白质的活性是必需的。 3.亚基的聚合: 有许多蛋白质是由二个以上亚基构成的,这就需这些多肽链通过非共价键聚合成多聚体才能表现生物活性。例如成人血红蛋白由两条α链,两条β链及四分子血红素所组成,大致过程如下:α链在多聚核糖体合成后自行释下,并与尚未从多聚核糖体上释下的β链相连,然后一并从多聚核糖体上脱下来,变成α、β二聚体。此二聚体再与线粒体内生成的两个血红素结合,最后形成一个由四条肽链和四个血红素构成的有功能的血红蛋白分子。 4.水介断链: 一般真核细胞中一个基因对应一个mRNA,一个mRNA对应一条多肽链,但也有少数的情况,即一种三思而行翻译后的多肽链经水介后产生几种不同的蛋白质或多肽。例如哺乳动物的鸦片样促黑皮激素原初翻译产物为265个氨基酸,它在脑下垂体前叶细胞中,POMC初切割成为N-端片断和C端片段的β-促脂解激素。然后N端片段又被切割成较小的N端片断和工9肽的促肾上腺皮质激素。而在脑下垂体中叶细胞中,β-促脂解激素再次被切割产生β-内啡肽;ACTH也被切割产生13肽的促黑激素(α-melanotropin)(图18-21)。
图18-21 POMC作为多种活性物质的前体 第一行为POMC前体,K、R为赖氨酸和精氨酸残基
…… 第四节 RNA的复制 小结 第十八章 蛋白质的生物合成(The Biosynthesis of protein) 第一节 参与蛋白质生物合成的物质 第二节 蛋白质生物合成过程 第三节 蛋白质合成后的分泌及加工修饰(当前页) 第四节 蛋白质合成的抑制剂 小结 第十九章 基因表达调控(Gene Expression and Its Regulation) 第一节 基因表达调控的现象和概念 …… |