|
《组织学与胚胎学》 > 第七章 神经组织
二、突触突触(synapse)是神经元传递登记处的重要结构,它是神经元与神经元之间,或神经元与非神经细胞之间的一种特化的细胞连接,通过它的传递作用实现细胞与细胞之间的通讯。在神经元之间的连接中,最常见是一个神经元的轴突终末与另一个神经元的树突、树突棘或胞体连接,分别构成轴-树(axodendritic)、轴-棘(axospinous)、轴-体(axosomatic)突触(图7-4)。此外还有轴-轴(axoaxonal)和树-树(dendrodendritic)突触等。突触可分为化学突触(chemical synapse)和电突触(electrical synapse)两大类。前者是以化学物质(神经递质)作为通讯的媒介,后者是亦即缝隙连接,是以电流(电讯号)传递信息。哺乳动特神经系统以化学突触占大多数,通常所说的突触是指化学突触而言。 突触的结构可分突触前成分(presynaptic element)、突触间隙(synaptic cleft)和突触后成分(postsynaptic element)三部分。突触前、后成分彼此相对的细胞膜分别称为突触前膜和突触后膜(presynaptic and postsynapticmembrane),两者之间在宽约15~30nm的狭窄间隙为突触间隙,内含糖蛋白和一些细丝。突触前成分通常是神经元的轴突终末,呈球状膨大,它们在银染色标本中呈现为棕黑色的环扣状,附着在另一神经元的胞体或树突上,称突触扣结(synaptic bouton)(图7-9)。
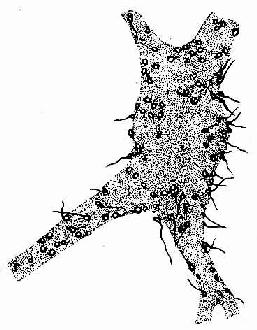
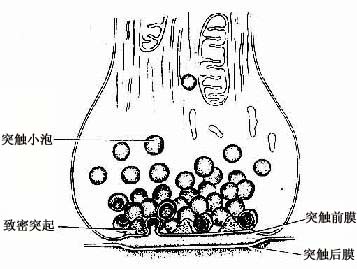
图7-9 脊髓运动神经元银染,示突触扣结 电镜下,突触扣结内含许多突触小泡(synapse vesicle),还有少量线粒体、滑面内质网、微管和微丝等(图7-10,7-11)。突触小泡的大小和形状不一,多为圆形,直径40~60nm,亦有的呈扁平形。突触小泡有的清亮,有的含有致密核芯(颗粒型小泡),大的颗粒型小泡直径可达200nm(图7-4)。突触小泡内含神经递质或神经调质。突触前膜和后膜均比一般细胞膜略厚,这是由于其胞质面附有一些致密物质所致(图7-10)。在突触前膜还有电子密度高的锥形致密突起(dense projection)突入胞质内,突起间容纳突触小泡。突触小泡表面附有突触小泡相关蛋白,称突触素Ⅰ(synapsinⅠ),它使突触小泡集合并附在细胞骨架上。突触前膜上富含电位门控通道,突触后膜上则富含受体及化学门控通道。当神经冲动沿轴膜传至轴突终末时,即触发突触前膜上的电位门控钙通道开放,细胞外的Ca2+进入突触前成分,在ATP的参与下使突触素I发生磷酸化,促使突触小泡移附在突触前膜上,通过出胞作用释放小泡内的神经递质到突触间隙内。其中部分神经递质与突触后膜上相应受体结合,引起与受体偶联的化学门控通道开放,使相应离子进出,从而改变突触后膜两侧离子的分布状况,出现兴奋或抑制性变化,进而影响突触后神经元(或非神经细胞)的活动。使突触后膜发生兴奋的突触称兴奋性突触(excitatory synapse),使突触后膜发生抑制的称抑制性突触(inhibitory synapse)。突触的兴奋或抑制,取决于神经递质及其受体的种类。
图7-10 化学突触超微结构模式图
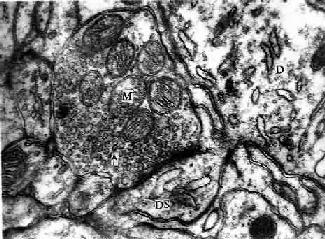
图7-11 豚鼠 小脑电镜像示突触 D树突,DS树突棘,M线粒体,↑突触小泡 (白求恩医科大学尹昕、朱秀雄供图) 一个神经元既可与其他神经元建立许多突触连接,亦可接受来自其他神经元的许多突触信息。一个神经元上突触数目的多少视不同的神经元而有很大差别,例如小脑的颗粒细胞只有几个突触,一个运动神经元要有1万个左右突触,而小脑的蒲肯野细胞树突上的突触就有10万个以上。一个神经元上众多的突触中,有些是兴奋性的,有些则是抑制性的。如果所有兴奋性突触活动的总和超过抑制性突触活动的总和,并足以刺激该神经元的轴突起始段产生动作电位时,则该神经元发生兴奋;反之,则表现为抑制。 根据突触前、后膜胞质面致密物质厚度的差异,可把突触分为Ⅰ、Ⅱ两型。Ⅰ型突触的突触后膜胞质面附有的致密物质较突触前膜的厚,两者不对称,突触间隙也较宽(30nm),故称不对称突触。Ⅱ型突触前、后膜的致密物质少,厚度相近,突触间隙较窄(20nm),称对称突触。有人认为Ⅰ型属兴奋性突触,Ⅱ型属抑制性突触。 神经递质的种类很多,包括:乙酰胆碱(acetylcholine,Ach);单胺类,如去甲肾上腺素(norepinephrine),多巴胺(dopamine,DA)和5-羟色胺(5-hydroxytryptamine,5-HT);氨基酸类,如γ-氨基丁酸(γ-aminobutyric acid GABA)、甘氨酸(glycine)、谷氨酸(glutamic acid)等。新近又发现大量的神经肽(neuropeptide),如P物质(substance P)、脑啡肽(enkephalin)、神经隆压素(neurotensin)、血管活性肠肽(vasoactive intestinal polypeptide,VIP)、胆囊收缩素(cholesystokinin)、加压素(vasopressin)、和下丘脑释放激素(hypothalamic releasing hormones)等约40多种。有些神经肽亦见于胃肠管的内分泌细胞,故总称为脑肠肽(braingut peptide)。这些肽类物质能改变神经元对经典神经递质的反应,起修饰经典神经递质的作用,故称为神经调质(neuromodulator)。不同形态大小的突触小泡所含的神经递质也不同,如圆形清亮小泡多数含乙酰胆碱,小颗粒型小泡含单胺类,大颗粒型小泡往往含神经肽。过去认为一个神经元一般只产生和释放一种神经递质,但近来应用免疫细胞化学法研究发现,某些神经元可产生和释放两种或两种以上的神经递质和(或)神经调质,其中一种往往是经典神经递质(Ach或NE),另一种则是神经肽。神经递质或神经调质共存的生理意义尚待研究。 突触后膜上的受体是一种膜蛋白,它能与相应的神经递质的结合而使突触后膜产生兴奋或抑制。神经递质的种类很多,受体的种类相应也很多。虽然一种受体只与相应的一种神经递质结合,但一种神经递质却可有不止一种受体。如乙酰胆碱受体就有N型(兴奋型)和M型(多数为兴奋型,少数为抑制型),去甲肾上腺素受体亦有α和β两类。所以,突触的兴奋或抑制,不仅取决于神经递质的种类(如γ-氨基丁酸是脑内一种抑制性神经递质),更重要的还取决于受体的类型。同一种神经递质在神经系统的不同部位有兴奋或抑制的不同效应,主要原因是突触后膜上受体类型的不同。突触后膜上的受体可直接与通道蛋白偶联或其本身就是一种通道蛋白,故当神经递质与这种形式的受体结合后使突触后膜产生的兴奋或抑制性变化十分迅速,所以把这种形式的受体称为快速作用受体,如乙酰胆碱N型受体属于快速作用受体。另外亦有慢速作用受体,这种受体一般与G蛋白偶联,再经过细胞内第二信使(环腺苷酸,cAMP)及蛋白磷酸化产生效应,它的作用比前者缓慢,但能把递质-受体相互作用所提供的微弱信号放大数千倍,如去甲肾上腺素β受体属于这种慢速作用受体。 释放到突触间隙的递质分子与突触后膜的受体结合产生生理效应后,很快便被相应的酶灭活(如Ach),或吸收入突触终末内被分解(如NE),以迅速消除该递质的作用,这样才能保证突触传递的灵敏性。递质的分解产物可被重新利用合成新的递质。非肽类递质除在胞体合成外,通常也可在轴突终末内合成,而肽类递质则只能在胞体内合成,释放后亦不能回收。合成的递质分子一般都贮存在突触小泡内。
|