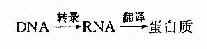|
《生物化学与分子生物学》 > 第十五章 核酸的结构与功能The Structure and Function of Nucleic Acid
第五节 RNA的结构与功能DNA是遗传信息的载体,遗传信息的作用通常由蛋白质的功能来实现,但DNA并非蛋白质合成的直接模板,合成蛋白质的模板是RNA。正常细胞遗传信息的流向是:
与DNA相比,RNA种类繁多,分子量相对较小,一般以单股链存在,但可以有局部二级结构,其碱基组成特点是含有尿嘧啶(uridin,U)而不含胸腺嘧啶,碱基配对发生于C和G与U和A之间,RNA碱基组成之间无一定的比例关系,且稀有碱基较多。此外,tRNA还具有明确的三级结构。 表15-7 RNA的分类
注:原核细胞不含后3种RNA
(一)信使RNA(mRNA)与不均一核RNA(hnRNA)遗传信息从DNA分子抄录到RNA分子中的过程称为转录(transcription)。在真核生物中,最初转录生成的RNA称为不均一核RNA(heterogeneous nuclearRNA,hnRNA),然而在细胞浆中起作用,作为蛋白质的氨基酸序列合成模板的是mRNA(messengerRNA)。hnRNA是mRNA的未成熟前体。两者之间的差别主要有两点:一是hnRNA核苷酸链中的一些片段将不出现于相应的mRNA中,这些片段称为内含子(intron),而那些保留于mRNA中的片段称为外显子(exon)。也就是说,hnRNA在转变为mRNA的过程中经过剪接,被去掉了一些片段,余下的片段被重新连接在一起;二是mRNA的5′末端被加上一个m7pGppp帽子,在mRNA3′末端多了一个多聚腺苷酸(polyA)尾巴。mRNA从5′末端到3′末端的结构依次是5′帽子结构,5′末端非编码区,决定多肽氨基酸序列的编码区,3′末端非编码区,和多聚腺苷酸尾巴。多聚腺苷酸尾一般由数十个至一百几十个腺苷酸连接而成。随着mRNA存在时间的延续,这段聚A尾巴慢慢变短。因此,目前认为这种3′末端结构可能与增加转录活性以及使mRNA趋于相对稳定有关。原核生物的mRNA没有这种首、尾结构。
图15-13 hnRNA与mRNA的结构比较 (涂斜线者为外显子,空白者为内含子) 1961年,Jacob和Monod首先提出了mRNA的概念。在真核细胞中,由于蛋白质是在胞浆中而不是在核内合成,因此显然要求有一个中间物将DNA上的遗传信息传递至胞浆中。后来的研究证实,这种中间物即信使RNA。mRNA的核苷酸序列与DNA序列相应,决定着合成蛋白质的氨基酸序列。它如何指导氨基酸以正确的顺序连接起来呢?不同的mRNA碱基组成和排列顺序都不同,但都只有A,G,C,U4种碱基。如果一个碱基就可以决定一个氨基酸,则只有四种变化方式,如果两个碱基决定一个氨基酸,则只有16种变化方式,都不能满足20种氨基酸的需要。1961年Crick和Brenner的实验得出了三个核苷酸编码一个氨基酸的结论,并将这种三位一体的核苷酸编码称做遗传密码(genetic code)或三联体密码,这样就可以有64种不同的密码,但此情况下必须假定有一些氨基酸使用两个以上的密码。这一假定很快就被证明是对的。遗传密码具有下列特征: (1)三个核苷酸组成一个密码子,每个密码子由三个前后相联的核苷酸组成,一个密码子只为一种氨基酸编码。共有64个密码子; (2)密码子之间不重叠使用核苷酸,也无核苷酸间隔; (3)一种氨基酸可有多个密码子,这个特点称为密码子的简并性; (4)密码子的通用性,所有生物从最低等的病毒直至人类,蛋白质合成都使用同一套密码子表(表15-8),仅有极少的例外,如特殊细胞器线粒体,叶绿体所用的密码稍有不同。(表15-9)。 表15-8 通用遗传密码及相应的氨基酸
表15-9 通用遗传密码与线粒体遗传密码之间的一些差异
注:下标横线者为与通用编码不同的编码 究竟哪一个密码子为哪一种氨基酸编码,即密码子与氨基酸之间的对应关系已在60年代研究解决了。1964年Nirenberg用一种RNA聚合酶体外合成了多聚尿苷酸、多聚腺苷酸等多聚核苷酸,将这些多聚核苷酸分别用于蛋白质的体外合成。发现,当所用的多聚核苷酸为多聚尿苷酸时,只有多聚苯丙氨酸合成,这意味着UUU为苯丙氨酸编码;用其它多聚核苷酸进行相应的实验后发现,CCC为脯氨酸编码,而AAA为赖氨酸编码;其后,有人又用核苷酸比例为已知,但是核苷酸序列随机的多聚核苷酸,以及用已知序列的含两种或两种以上核苷酸的多聚核苷酸进行相应的实验,将结果加以数理统计处理,又解读了一批密码子,其中包括三个终止码,最后,还有一些密码子是通过合成已知序列的三聚核苷酸与核蛋白体和载有放射性同位素标记的氨基酸的tRNA共沉淀原理予以解读的。在所有密码子中,AUG不仅为蛋氨酸编码,而且又是翻译(translation,以mRNA上的遗传信息指导核蛋白体上多肽链合成的过程)的起始信号,UAA、UAG和UGA不为任何氨基酸编码,而是作为翻译的终止信号,统称为终止码(stop codon),又常被叫作无意义码(nonsense codon)。 大多数氨基酸是由一个以上的密码子所编码。这个事实提出了一个问题:编码同一种氨基酸的一组密码子的使用频率是否都相同?细致的分析表明,无论是原核生物,还是高等真核生物,密码子的使用频率并不是平均的,有些密码子的使用率很高,有些则几乎不使用,其使用频率主要与细胞内tRNA含量呈正相关。
(二)转运RNA(tRNA)tRNA(transfer RNA)是蛋白质合成中的接合器分子。tRNA分子有100多种,各可携带一种氨基酸,将其转运到核蛋白体上,供蛋白质合成使用。tRNA是细胞内分子量最小的一类核酸,由70~120核苷酸构成,各种tRNA无论在一级结构上,还是在二、三级结构上均有一些共同特点。tRNA中含有10%~20%的稀有碱基(rare bases),如:甲基化的嘌呤mG、mA,双氢尿嘧啶(DHU)、次黄嘌呤等等。此外,tRNA内还含有一些稀有核苷,如:胸腺嘧啶核糖核苷,假尿嘧啶核苷(Ψ,pseudouridine)等。胸腺嘧啶一般存在于DNA中;在假尿嘧啶核苷中,不是通常嘧啶环中1位氮原子,而是嘧啶环中的5位碳原子与戊糖的1′位碳原子之间形成糖苷键。
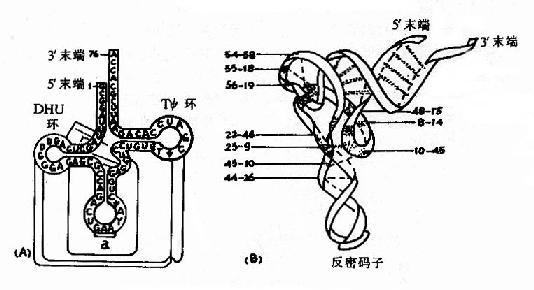
tRNA分子内的核苷酸通过碱基互补配对形成多处局部双螺旋结构,未成双螺旋的区带构成所谓的环和襻。现发现的所有tRNA均可呈现图15-14所示的这种所谓的三叶草样(clover leafpattern)二级结构。在此结构中,从5′末端起的第一个环是DHU环,以含二氢尿嘧啶为特征;第二个环为反密码子环,其环中部的三个碱基可以与mRNA中的三联体密码子形成碱基互补配对,构成所谓的反密码子(anticodon),在蛋白质合成中起解读密码子,把正确的氨基酸引入合成位点的作用;第三个环为TΨ环,以含胸腺核苷和假尿苷为特征;在反密码子环与TΨ环之间,往往存在一个襻,由数个乃至二十余个核苷酸组成,所有tRNA3′末端均有相同的CCA-OH结构,tRNA所转运的氨基酸就连接在此末端上。(图15-14A)
图15-14 tRNA的二级与三级结构 A.二级结构(a示反密码环及反密码) B.三级结构(数字示可能的非常见核苷酸对相互作用) 通过X-射线衍射等结构分析方法,发现tRNA的共同三级结构均呈倒L形(图15-14B),其中3′末端含CCAOH的氨基酸臂位于一端,反密码子环位于另一端,DHU环和TΨ环虽在二级结构上各处一方,但在三级结构上却相互邻近。tRNA三级结构的维系主要是依赖核苷酸之间形成的各种氢键。各种tRNA分子的核苷酸序列和长度相差较大,但其三级结构均相似,提示这种空间结构与tRNA的功能有密切关系。
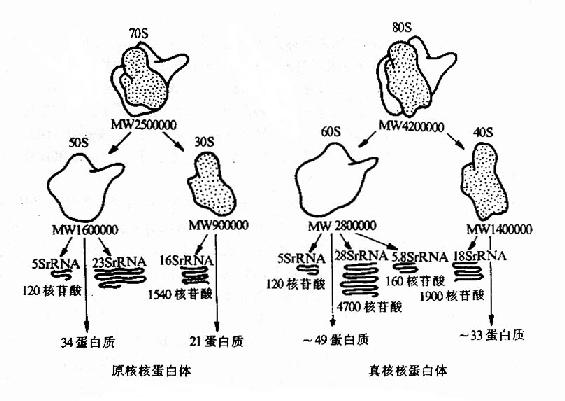
(三)核蛋白体RNA(rRNA)核蛋白体RNA(ribosomalRNA)是细胞内含量最多的RNA,约占RNA总量的80%以上,是蛋白质合成机器枣核蛋白体(核糖体)(ribosome)的组成成分。核糖体蛋白(ribosmal protein,rp)有数十种,大多是分子量不大的多肽类,分布在核蛋白体大亚基的蛋白称为rpl,在小亚基的称rps。 原核生物和真核生物的核蛋白体均由易于解聚的大、小亚基组成。对大肠杆菌核蛋白体的研究发现其质量中三分之二是rNRA,三分之一是蛋白质。rRNA分为5S、16S、23S三种。S是大分子物质在超速离心沉降中的一个物理学单位,可反映分子量的大小。小亚基由16SrRNA和21种rps构成,大亚基由5S、23s rRNA和31种 rpl构成。真核生物核蛋白体小业基含18S rRNA和30多种rps,大亚基含28S、5.8S、5S三种rRNA,近50种rpl。各种生物核蛋白体小亚基中的rRNA具有相似的二级结构(图15-15)。
图15-15 原核生物与真核生物核蛋白体的结构比较 (线粒体核蛋白体的结构与原核相似) 无论在试管内或细胞内,大、小亚基都易于组成核蛋白体整体或分离成两部分。几十种多肽是如何互相联结,又怎样与几种rRNA相连的呢?用提纯了的亚基所有的肽和rRNA在试管内混合,发现不需加入酶或ATP就可以自动组装成为有活性的亚基,但rRNA之间却不能互相替代,也即说这种自我组装过程是以rRNA为主导的。虽然所有多肽在组装中也是缺一不可的,但不同的肽可能有酶的作用或起别构效应。现已证明某些核糖体蛋白具有酶的功能,但基中大多数还未弄清其具体作用。
(四)其它RNA分子小核RNA(snRNA,smallnuclear RNA)存在于真核细胞的细胞核内,是一类称为小核核蛋白体复合体(snRNP)的组成成分,有U1,U2,U4,U5,U6snRNA等,均为小分子核糖核酸,长约106?89个核苷酸,其功能是在hnRNA成熟转变为mRNA的过程中,参与RNA的剪接,并且在将mRNA从细胞核运到细胞浆的过程中起着十分重要的作用(表15-10)。 表15-10 snRNA的种类与功能
小胞浆RNA(scRNA,smallcytosol RNA)又称为7SLRNA,长约300个核苷酸,主要存在于细胞浆中,是蛋白质定位合成于粗面内质网上所需的信号识别体(signal recognization particle)的组成成分。
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||