|
《实用免疫细胞与核酸》 > 第二十三章 DNA重组技术及其在基因诊断中的应用
第二节 分子克隆载体载体(vector)是指运载外源DNA有效进入受体细胞内的工具。载体同外源DNA在体外重组成DNA重组分子,在进入受体后形成一个复制子,即形成在细胞内能独自进行自我复制的遗传因子。因此,作为载体应该满足以下几方面的要求:①有某种限制酶的一个切点,最好是有许多种限制酶的切点,而且每种酶的切点只有一个;②外源DNA插入后不影响载体在受体细胞中进行自我复制,载体应对受体细胞无害,以及载体能接纳尽可能大的外源DNA片段;③有利于选择的标记基因,可以很方便地知道外源DNA已经插入,以及把接受了载体的受体细胞选出;④具有促进外源DNA表达的调控区。 重组DNA技术中最常用的载体有质粒、噬菌体λ,柯斯质粒(cosmid)和噬菌体M13。它们的受体细胞都是大肠杆菌。这四种载体的大小和结构尽管各不相同,但它们的共同特点是:①都能在大肠杆菌中自主复制,而且能连同所带的外源DNA一起复制;②都很容易同细菌DNA分开并加以纯化;③都有一段DNA对于它们自身在细菌中的增殖不是必需的。因此,外源DNA可以插入这一段DNA中,或是置换这一段DNA而不影响载体的复制。根据这一特点,载体又可分成插入型和置换型两大类。
一、质粒质粒(plasmid)是许多种细菌中发现的染色体外的遗传因子,它是闭合环状的双链DNA分子,大小从1kb直到200kb以上。质粒所带的基因通常有利于宿主细胞。例如基因的产物是某些抗生素或对某些抗生素的抗性、能降解某些复杂的有机化合物、产生大肠杆菌素和肠毒素、产生限制酶或修饰酶等。 在天然条件下,许多种质粒是通过类似于细菌接合的过程传递给新的宿主。在实验室条件下,质粒则可通过转化过程进入受体细胞。此时,受体细胞已经过处理而处于感受态,即它的细胞暂时易于让小的DNA分子透过。受体细胞由于质粒的进入而产生了新的表型。如对某种抗生素的抗性,这样就可用这种抗生素作为选择条件,选出被质粒转化的受体细胞。 质粒复制时利用宿主细胞复制自身染色体的同一组酶系。有些质粒处于“严紧控制”之下,即它们的复制是同宿主细胞的复制偶联同步的。因此在每个细胞中的质粒只有一份拷贝,或最多只有几份拷贝。这称为“严紧型”质粒。另一些质粒则是处于“松弛控制”之下的“松弛型”质粒。每个细菌中可以有10~200份拷贝。更重要的是松弛型质粒的拷贝数,可因宿主细胞停止合成蛋白质而增加至几千份。宿主细胞不合成蛋白质时,染色体和严紧型质粒的复制都中止。但松弛型质粒仍继续进行复制。因此在增殖松弛型质粒时,总是要让宿主菌经氯霉素处理(在培养液中加入适量的氯霉素)抑制蛋白质的合成。
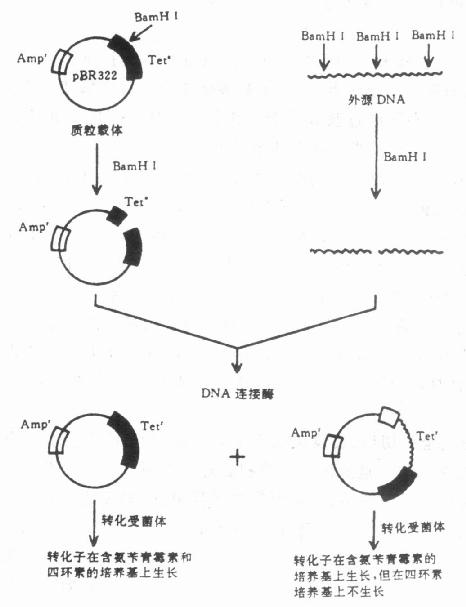
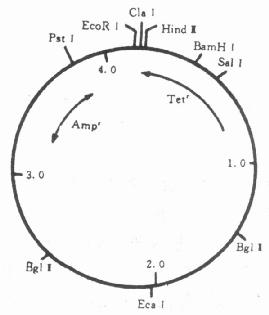
图23-3 pBR322的基本结构 (一)大肠杆菌质粒应用的最广泛的质粒载体是pBR322,它属松弛型质粒,有抗氨苄青霉素和抗四环素两个抗性基因可作为标记基因,有许多种常用的限制酶的切点。它全长4352个核苷酸,排列顺序已全部测定,基本结构如图23-3。 当需要用pBR322来克隆BamHI酶切的一个DNA片段时,一般的做法是如图23-4所示。
图23-4 pBR322克隆DNA片段的程序 从图23-4可见,外源DNA片段同pBR322都经Bam HⅠ酶切,所以有相同的单链“粘性末端”,通过DNA连接酶可以构成重组DNA分子。由于pBR322的BamHⅠ酶切点在四环素的抗性基因中,所以Bam HⅠ酶切后使这个抗性基因失活。如果酶切后的质粒pBR322自身又重新连接,则恢复对四环素的抗性,转化子就仍然具有对二种抗生素的抗性。如果pBR322中插入外源DNA,则转化子失去了四环素抗性表型,而只有对氨苄青霉素的抗性,这样就可根据转化子的抗性而把重组质粒转化的细菌选出,然后扩增细菌,大量回收质粒DNA,从而达到大量扩增外源DNA的目的。 (二)真核生物中的质粒真核生物中的酵母是国内外广泛研究的载体-受体系统之一。这是一个共价闭环DNA分子(cccDNA),平均周长2μm,所以称为2μm DNA质粒,在每个酵母细胞里约有100个拷贝。已有人把2μm DNA同pBR322 DNA构建成双功能质粒,可在大肠杆菌和酵母菌中复制。这种质粒带有啤酒酵母的二个基因(his 3和Leu 2),所以很容易用酵母的营养缺陷型菌株来选出。
二、噬菌体和病毒载体噬菌体λ是最主要的一种载体。自从1974年以来,已用野生型λ改造和构建出一系列噬菌体载体。 噬菌体λ是温和噬菌体。基因组是长约50kb的双链DNA分子。在噬菌体λ颗粒中,DNA是线状双链分子带有单链的互补末端。末端长12个核苷酸,这称为粘性末端,简写COS。当噬菌体感染宿主细胞后,双链DNA分子通过COS而成环状。在感染早期,环状DNA分子进行转录。在此期间,噬菌体有两条复制途径可供选择:一是裂解生长。环状DNA分子在宿主细胞里复制若干次,合成了大量的噬菌体基因产物,形成子代噬菌体颗粒,成熟后使细菌裂解,释放出许多新的有感染能力的病毒颗粒。另一是溶源性生长。噬菌体DNA整合进宿主菌的基因组,然后象细菌染色体上的基因一样进行复制,并传递给下一代细菌。 (一)噬菌体λ载体的构建野生型噬菌体λDNA全长约50kb,上有65种限制酶酶切点,除ApaⅠ、NaeⅠ、NarⅠ、NheⅠ、Sna BⅠ、XbaⅠ和XhoⅠ等7种限制酶各有一个切点外,其余都多于2个。有些酶切点在λ增殖所必需的基因区域内。因此,噬菌体λ必须经过改造才能用作载体。现在用的λ载体大都除去了某种限制酶的酶切点。因此,作为载体的噬菌体λ都短于野生型。 噬菌体λ载体有两种类型:①插入型。由于改建后的噬菌体λDNA都短于野生型,所以可插入外源DNA。只要插入的位置不影响噬菌体增殖。而且噬菌体DNA缺失越长,插入片段就可越大。②置换型。噬菌体λ的基因组可分为三个区域,左侧区包括使噬菌体DNA成为一个成熟的、有外壳的病毒颗粒所需的全部基因,全长约20kb。右侧区内包含所有的调控因子、与DNA复制及裂解宿主菌有关的基因,这个区域约长12kb。中间区域约长18kb,这一段DNA可以被外源置换而不会影响噬菌体λ裂解生长的能力。置换型噬菌体λ是使用最广泛的载体。 噬菌体λ裂解生长的能力同包装在头蛋白中的λDNA的大小有关。当DNA的长度短于野生型的78%或超过105%时,噬菌体的活性就急剧下降。因此要求λ载体DNA和外源DNA长度之和在39~53kb之间。 置换型载体DNA用选定的限制酶完全酶切后,要设法除去中间片段,只留下左臂和右臂以便用外源DNA片段连接,包装成重组噬噬体。这是因为左臂和右臂与中间片段间都有单链“粘性末端”,在连接酶作用下可以重新恢复原来的结构,从而影响了同外源DNA的连接。区分重组噬菌体形成的噬菌斑和重新恢复的载体噬菌体形成的噬菌斑的方法是用Xgal蓝色噬菌斑试验。有些噬菌体载体的中间片段带有编码β半乳糖苷酸的基因。当这种噬菌体感染lac宿主细胞并在含有Xgal(5溴4氯3吲哚βD半乳糖苷)的培养基上生长时,半乳糖苷酸与Xgal反应的产物为不溶性的靛蓝染料。因此,含有中间片段的重新恢复的噬菌体形成蓝色噬菌斑;而同外源DNA连接的重组噬菌体形成的噬菌斑是无色透明的。 (二)单链噬菌体载体最常用的单链噬菌体载体是M13和它的改建噬菌体。M13是丝状噬菌体,有长约6500个核苷酸的闭环DNA基因组。M13附着在大肠杆菌的F性菌毛上,所以它们只能感染雄性细菌,即F’或Hfr细菌。当噬菌体进入细菌细胞后,单链的噬菌体DNA转变成双链复制型(RF)。从细胞中分离的RF可用作克隆双链DNA的载体。当每个细菌细胞里积聚了200到300份RF型拷贝时,M13开始不对称的合成,即只大量合成DNA双链中的一条链。单链DNA掺入成熟的噬菌体颗粒,颗粒不断地从感染细胞上芽生。M13感染虽不杀死细胞,但细胞的生长受到一定的抑制,所以形成混浊的噬菌斑。 M13的基因组中除有一段507个核苷酸,其余都含有与其复制有关的遗传信息。因此,只有在这一段中可插入外源DNA而不致影响噬菌体的活性。用M13作为分子克隆载体的优点是从细菌中释放出来的噬菌体颗粒只含单链DNA,其中包含了克隆DNA片段的二条互补链中的一条,因此可用来作为对DNA测序的模板。另外,可用来产生单链的DNA探针以选择和分离互补的RNA。M13的RF型是双链DNA,便于限制酶的识别和切割,克隆进双链的外源DNA。从细菌培养液中大量抽取单链DNA。问题是插入M13的外源DNA超1000bp时就不稳定,在噬菌体增殖时会出现缺失。一般说,插入300~400bp是相当稳定的。 (三)猴病毒40(SV40)猴病毒40(SV40)的基因组常用来作为克隆载体,把外源DNA转入哺乳类细胞。猴病毒40是球形动物病毒,直径40nm,呈20面体,有一个共价闭环的双链DNA基因组,全长5244bp。它在猴肾中增殖。被SV40感染的细胞都会出现SV40的T抗原。早已证明完整的小鼠染色体β珠蛋白基因(包括所有的间插顺序和两侧顺序)整合在SV40中后,在被感染的猴肾细胞中都能正确地转录和翻译。表明猴肾细胞的拼接系统能有效地处理小鼠β珠蛋白的初级转录本。 可是,SV40作为克隆载体有其局限性:①SV40基因组的晚期功能区的裂解性,不利于外源DNA的重组和表达;②早期功能区与病毒的致癌性有关,使用时有顾虑;③重组DNA片段的大小受体限制。 (四)逆转录病毒逆转录病毒是RNA病毒,它有三个基因:gag-编码病毒的核心蛋白;pol-编码逆转录酶;env-编码病毒的被膜糖蛋白。有的逆转录病毒还带有癌基因(vonc),即有的逆转录病毒有致癌作用。近年来,已设计构建成一些缺陷型病毒(defective virus)使逆转录病毒成为有用的基因载体,成功地把抗药性基因转入了人体造血前体细胞,并在细胞中表达。Hock等选用了缺失编码病毒外壳蛋白基因的逆转录病毒,因此不能合成自身的外壳,但它有识别外壳蛋白进行包装的信号(一段尚未鉴定的DNA顺序)。用这种缺陷的逆转录病毒去感染某种细胞株,这种细胞株包含有辅助病毒(helpervirus)。辅助病毒能合成蛋白外壳,但缺失了识别蛋白外壳进行包装的信号,因此它不能包装成病毒颗粒。当用逆转录病毒感染细胞株后,逆转录病毒的RNA进入辅助病毒的外壳蛋白,成为病毒颗粒。这时把受感染的细胞同骨髓细胞一起培养,包装在辅助病毒外壳蛋白中的逆转录病毒RNA,进入骨髓细胞,病毒DNA插入宿主细胞基因组,基因的活性得到表达。这时,由于骨髓细胞里面没有辅助病毒,所以整合进宿主基因组的逆转录病毒,不再有外壳蛋白可供包装,因此也就无法增殖,而只能被“陷”在宿主基因组中,通过细胞分裂而传给下一代子细胞。
三、柯斯质粒1978年Collins和Hohn构建一种新型的大肠杆菌克隆载体,命名为cosmid(柯斯质粒)。它是用正常的质粒同噬菌体λ的cos位点构成。例如,柯斯质粒pHC79系由质粒pBR322和噬菌体λ的cos位点的一段DNA构成(图23-5),全长43.kb。在包装时,cos位点打开而产生噬菌体λ的粘性末端。由于pHC79有pBR322DNA,所以也就有氨苄青霉素抗性和四环素抗性两个标记。图23-5说明了克隆基因时有用的限制酶切点。所以可以说柯斯质粒是一类特殊的重组质粒。
图23-5 柯斯质粒pHC79 设计构建的柯斯质粒一般长4~6kb。其上的cos位点的一个重要作用是识别噬菌体的外壳蛋白。凡具有cos位点的任何DNA分子只要在长度相当于噬菌体基因组,就可以同外壳蛋白结合而被包装成类似噬菌体λ的颗粒。因此,插入柯斯质粒的外源DNA可大于40kb。重组的柯斯质粒可象噬菌体λ一样感染大肠杆菌,并在细菌细胞中复制。如把宿主菌在含氯霉素的培养基中生长,柯斯质粒可以扩增到宿主细胞DNA总量的50%左右。 采用柯斯质粒作载体的困难是:①载体自身只相当于可以插入片段的1/10左右,因此往往会出现载体同载体自身连接,结果在一个重组分子内可有几个柯斯质粒载体连在一起。但用碱性磷酸酶处理,可阻止载体分子自身连接;②大小不等的外源片段相互连接后插入同一个载体分子,结果使在基因组内本来不是相邻的片段错乱地连接成一个片段,会影响实验结果的分析,后来专门选出30~45kb的外源DNA插入载体DNA,此时,每个载体只可能插入一个外源片段,因为如果二处片段,则将超过包装成噬菌体颗粒的限度;③细菌的菌落体积远大于噬菌斑,因此如用柯斯质粒制备基因文库,则筛选所需的含某一DNA片段的菌落很费时间。现虽建立了高密度菌落筛选法,但由于柯斯质粒制成的基因文库常常不太稳定,插入的大片段外源DNA有可能通过同宿主基因组交换而致丢失等,所以最常使用的还是噬菌体载体。 现将上面提到的四种载体作一比较(表23-3) 表23-3 四种载体的比较
* 外源DNA如超过10kb,则重组质粒的转化率和DNA得率都非常低 ** 已有个别质料可用于此目的
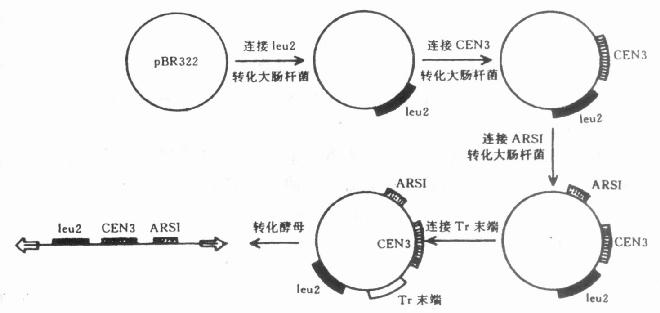
四、其他载体现在提到的分子载体有的不是用于克隆某基因,而是作为外源基因进入受体细胞的运载工具。 (一)转座子载体这里主要提一下有关果蝇转座因子(P因子)作为基因载体。因为这是很有希望的真核生物基因工程的载体,果蝇的P因子约长3kb,每端各有反向重复顺序,当它插入基因组时,可以激活或灭活近旁的基因,当它从基因组上切下来时,也可能把近旁的基因或DNA顺序一起切下而引起突变。现在已能把带有某种限制酶切点的P因子克隆到质粒pBR322中,然后在P因子酶切点上插入外源DNA。经过扩增后,将这种重组DNA用微量注射仪直接注入果蝇的受精卵中。结果所克隆的基因能随P因子插入受体细胞的基因里,并且得到表达。 (二)人工微小染色体包括人体在内的高等生物基因工程中遇到的一个难题是缺乏合适的基因载体。要求载体对受体细胞没有危害,能安全、稳定地转给后代,使基因的性状得到连续的表达。这对校正某种遗传缺陷或改进某种性状都是十分有用的。目前一些实验室正致力于构建人工微小染色体(microchromosome)。 构建染色体应包括:基因、复制起点、着丝粒和端粒。染色体上需带有基因是不言而喻的。复制起点是染色体复制所不可缺少的。着丝粒保证复制后的染色体均等地分配到两个子细胞中。端粒具有某种特定结构以保证染色体的个体性。 首先,构建的是酵母的人工微小染色体。天然的酵母染色体为150~1000kb。人工构建的微小染色体长55kb,在细胞内很稳定,只是每个细胞里的拷贝数很少。比如已构成的人工微小染色体为7~15kb,每个细胞里的拷贝数虽然增多,但不够稳定。 人工构建酵母微小染色体的具体步骤见图23-6。从图中可以看到,先用质粒pBR322为材料,连接酵母的标记基因Leu2(亮氨酸)后去转化大肠杆菌,然后再接上酵母染色体上分离得到的着丝粒CEN3,再去转化大肠杆菌以增殖重组质粒,取得下一步实验用的DNA。接下去是连接从酵母(真核生物)中分离得到的ARS1。ARS是指自主复制顺序(autonomouslyreplicat-ing sequence),这个顺序中可能包含着复制起点。目前已从非洲爪蟾的线粒DNA,衣藻和烟草叶绿体和核DNA中分离ARS。最后一个步骤是接上染色体端粒。用的是四膜虫(Te-trahymena)的rDNA末端(Tr末端)。这样构建成的质粒去转化酵母细胞后,在细胞内断开成为线状重组DNA分子,可以象天然染色体一样地复制和分离,这就是酵母线状质粒YLp4。
图23-6 构建酵母人工微小染色体的步骤
…… 第八节 PCR临床应用领域及特点 第九节 其它链扩增技术及应用范围 参考文献 第二十三章 DNA重组技术及其在基因诊断中的应用 第一节 工具酶 第二节 分子克隆载体(当前页) 第三节 DNA重组实验中常用的技术 第四节 遗传病与肿瘤的基因诊断 第五节 “DNA指纹”与法医鉴定 参考文献 …… |