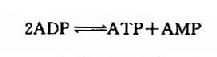|
《生物化学与分子生物学》 > 第十四章 神经组织生化(Biochemistry of Neural Tissue)
第二节 脑代谢的某些特点
一、能量供应(Energy Supply)脑的活动瞬息万变,需要大量能量的及时供应。脑细胞本身的生物高分子(核酸及蛋白质)的合成以及神经递质的合成与释放固然都是耗能的过程,但这些尚不足以说明为什么脑细胞的功能活动较之其他组织细胞要消耗更多的能量。脑的能量消耗主要在于经常不断地把Na+泵出细胞外,使去极化(depolarization)后的膜迅速恢复膜电位,以维持神经的兴奋和传导。脑的代谢率(metabolic rate)是很高的,它可以用单位时间的耗氧量(oxygenconsumption)和基质消耗量或产物生成量作指标来表示,如表14-1。 表14-1 正常青年男子的脑血流量和代谢率
脑血流量占心输出量的15%,耗氧量占全身总耗氧量(约250ml/分)的20%,然而脑的重量只不过占体重的2%。分析流入和流出脑组织血液的化学成分(动静脉差法)发现,除了葡萄糖外,其他可作为能源的物质没有明显的减少。由上表可见每百克脑组织每分钟产生的二氧化碳和消耗的氧均为3.5ml(156微克分子),呼吸商(respiratoryquotient)为1。再者,按化学计算(1克分子葡萄糖完全氧化要消耗6克分子的氧),葡萄糖的消耗率还稍高于氧的消耗率。 这些事实综合起来说明,脑组织是以葡萄糖的氧化来供能的,甚至可以说,至少在正常条件下,脑组织唯一利用糖作为能源。因为脑中糖原含量很少(小于0.1%),所以必须依赖血糖的供应。虽然脑组织还可以利用酮体,但必须以低血糖为前提,例如在饥饿引起酮血症(ketonemia)的情况下。如果血糖和血酮体均增高时(糖尿病酮血症),脑仍然优先利用葡萄糖以供能。有人认为,脑利用酮体作能源是对饥饿时低血糖的适应,长期慢性饥饿的病人,脑的耗氧量的一半可用来氧化酮体。 脑细胞含有完整的糖酵解(EMP)酶系,已糖激酶活性约为其他组织的20倍。但是即使最大程度地发挥糖酵解的作用也不能满足供能的需要,而必须依赖糖的有氧氧化。所以氧的供给一刻也不能中断。由于脑组织主要依赖糖的有氧氧化供给能量,所以它对缺糖和缺氧均极敏感。血糖下降50%即可致昏迷,而中断(流向脑的)血流几分钟就可引起死亡。临床上使用大剂量胰岛素做为治疗手段也可以引起昏迷。胰岛素对脑的这种影响,现在认为是间接作用的结果,也就是因为胰岛素降低了血糖之故。因为胰岛素是大分子物质,它不能透过血脑屏障,这和对其他组织或周围神经组织的作用不同。在那些组织,胰岛素的作用可能主要是直接影响其对葡萄糖的摄取。 脑内ATP的水平甚高,它的合成和利用均很迅速。据测定,脑内ATP末端磷酸基的半数更新时间平均只有约3秒钟,脑组织的磷酸肌酸(CP)水平比ATP还要高,它可看作是ATP末端高能磷酸键的一种贮存形式。在磷酸肌酸激酶(CPK)的催化下,ATP和CP可相互转变。
这是一个等能反应,因此在相互转变中不致于有能量的丢失。脑中CPK活性较强,有利于能量的贮存和动用。此外,脑组织中还有活性较强的肌激酶(myokinase),它可催化下列反应。此反应的意义在于“挖掘潜力”,即充分利用ATP中的两个高能磷酸键。
上述两种激酶都是维持脑内高水平ATP的有力保证。
二、类脂的组成和代谢除脂肪组织外,脑是全身含脂类最多的组织,但脂肪组织主要含甘油三酯(贮存脂),而脑组织中的脂类几乎全是类脂。脑干重的1/2是脂类,这是就全脑平均而言,如果分别测定脑灰质和脑白质的化学成分,就会发现灰质含水份和蛋白质较多,脂类仅占干重的1/3;而白质中的脂类含量较多,约占干重的55%。 表14-2 正常成人脑的脂类组成
由上表可见,以湿重计,脑白质中脂类含量约三倍于灰质。这种差别主要是由于白质中的神经纤维外被以髓鞘(myelin sheath),而髓鞘的脂类可高达干重的70-80%。 脑中的类脂主要用以构成神经元(neurone)的质膜和髓鞘。这些膜性结构与其他组织细胞的膜结构有共同之处,即都是由类脂与蛋白质构成的复合物,但在类脂的组成和代谢上亦有一些特点。尤其是髓鞘,它含有某些特殊的类脂成分,这些成分或者仅见于髓鞘,或者髓鞘中含量较多,而在其他组织中则较少见,例如缩醛磷脂(plasmalogen)和脑苷脂(cerebroside)。 脑不从血中摄取脂肪酸,本身也不含游离脂肪酸,那么组成这些类脂的脂肪酸是从哪里来的呢?实验证明,脑中的脂肪酸和胆固醇都可由乙酰CoA合成,而乙酰CoA的主要来源还是葡萄糖。 髓鞘形成(Myelination)之前的未成熟的脑组织含胆固醇和磷脂较多,而含脑苷脂极少,脑苷脂合成酶系的活性也极低。当髓鞘形成时,此酶系的活性升高,脑苷脂的含量亦相应增多,髓鞘形成与神经系统的发育和功能密切相关,而髓鞘脱落(demyelination)是神经系统疾病的重要的病理改变之一。髓鞘的代谢特点是正在进行髓鞘形成时代谢很快,一旦形成之后就变得很慢,成为体内最稳定的一种结构。据认为,这是由于髓鞘缺乏催化类脂分解代谢的酶系。已经形成的髓鞘,除了个别成分(如三磷酸肌醇磷脂)有较高的更新率外,其他磷脂和胆固醇等的更新率均甚低。
三、谷氨酸的代谢与功能脑的游离氨基酸组成与血浆有很明显的差别,这是由于血脑屏障的特点和脑本身氨基酸代谢特点造成的。脑和血浆中某些游离氨基酸的含量对比如表14-3。 表14-3 人脑和血浆中某些游离氨基酸的含量
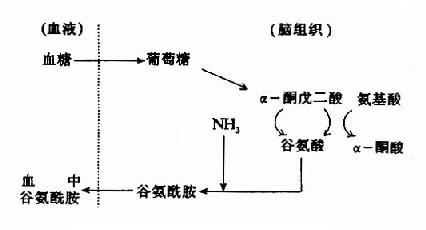
由表14-3可见,脑中游离氨基酸以谷氨酸(Glu)含量最高,它比其在血浆中的浓度要高出200倍以上。谷氨酸、谷氨酰胺(Gln)和γ氨基丁酸(GABA)三者含量总和约占脑中游离氨基酸总量的一半。所以,在脑的氨基酸代谢中,谷氨酸占有重要位置。 然而,谷氨酸难以通过血脑屏障,脑内谷氨酸来源于自身的合成,同位素示踪实验表明脑内谷氨酸合成的原料是葡萄糖,它来自血糖。葡萄糖进入脑细胞后先转变成α-酮戊二酸(α-KG),后者可在谷氨酸脱氢酶的催化下转变成谷氨酸,亦可经转氨基作用生成谷氨酸,一般认为后一途径更切合实际,因为谷氨酸脱氢酶(GDH)催化逆反应时KmNH4+为8mM,远高于细胞内氨之浓度。谷氨酸在谷氨酰胺合成酶的作用下与氨结合成为谷氨酰胺,这是一个耗能反应(消耗ATP),脑中谷氨酰胺合成酶的活性强,其KmNH4+仅为0.39mM。所生成的谷氨酰胺,与谷氨酸不同,可以通过血脑屏障而进入血中,这样,脑组织从血中摄入葡萄糖,通过代谢,还血液以谷氨酰胺,清除了脑中的氨,以免氨的积存危害脑的功能。
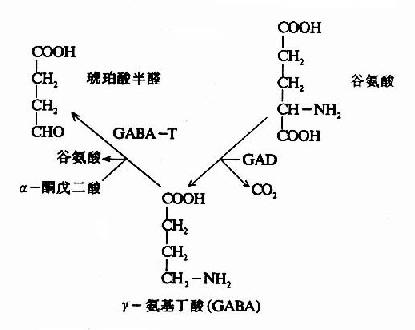
图14-1 脑中谷氨酸代谢与氨的消除 脑中谷氨酸代谢的另一个特点是脱羧生成γ-氨基丁酸(又名γ-氨酪酸GABA),催化此反应的酶是谷氨酸脱羧酶(GAD),它需要磷酸吡哆醛作辅酶。GABA是一种抑制性的神经递质,仅见于中枢神经系统。脑内GABA主要贮于灰质,特别是纹状体、黑质、小脑的齿状核等处。 GABA对中枢神经元有普遍性抑制作用。1963年曾有人提出,GABA能作用于突触前神经末梢,减少兴奋性递质的释放,从而引起抑制。这种效应称为突触前抑制(presynaptic inhibition)。GABA在脊髓中的作用就是以突触前抑制为主。在脑内则GABA主要是引起突触后抑制(postsynaptic inhibition)。睡眠时皮层释放GABA增多,因此有人认为GABA可能与睡眠、觉醒的生理机能有关。
在神经元胞体和突触(synapse)的线粒体内含有大量的γ-氨基丁酸转氨酶(GABA-T),它可催化GABA与α酮戊二酸之间的转氨作用,生成琥珀酸半醛(succinic acid semialdehyde)和谷氨酸。这可看作是GABA灭活的一种方式。GABA-T也是需要磷酸吡哆醛作辅酶,但与GAD比较,它同磷酸吡哆醛的亲和力大,所以当体内维生素B6缺乏时,主要影响GAD的活性。例如,使用异烟肼治疗结核病时,由于异烟肼能与维生素B6(吡哆醛)结合成异烟腙(isoniazone),加速维生素B6从尿中排泄,引起脑组织内维生素B6浓度下降,GAD活性亦下降,结果GABA的合成受阻,容易使中枢过度兴奋而发生抽搐等症状。所以长期使用异烟肼时应合并使用维生素B6。此外,临床上对于惊厥、妊娠呕吐的病人,也常使用维生素B6,其道理也是提高脑组织内GAD的活性,使GABA生成增多,中枢抑制相对加强。
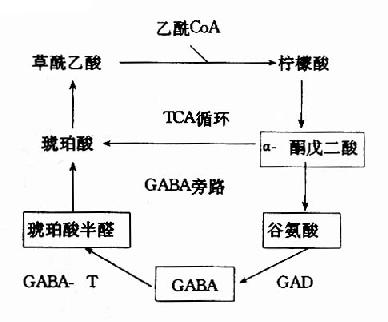
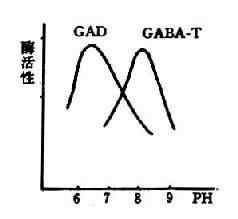
图14-3 GAD与GABA-T的作用最适pH GABA经转氨作用后的产物琥珀酸半醛可脱氢生成琥珀酸,后者进入三羧酸循环而被氧化利用。因此,与脑组织中的三羧酸循环相连系,存在着一条GABA代谢旁路(GABa shunt)。 谷氨酸脱羧酶与γ-氨基丁酸转氨酶的协同作用对保持脑中GABA一定浓度有重要意义。两种酶的最适pH不同,GAD的最适pH为6.5,而GABA-T则为pH8.2。(图14-3)由此可见,脑细胞内pH稍有变动就可明显改变这两种酶的活性对比。当酸中毒时,脑中GAD活性增强而GABA-T活性减弱,可致脑中GABA水平上升,呈现中枢抑制;反之,当碱中毒时脑中GABA-T活性增强而GAD活性减弱,脑中GABA水平下降,易于发生痉挛。 尚须指出,谷氨酸对神经中枢有兴奋作用,而其脱羧产物GABA却有抑制作用,所以谷氨酸的代谢与中枢的兴奋和抑制调节有关。此外,通过GABA代谢旁路,也把脑的氧化代谢与兴奋抑制功能联系起来了。
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||